Molecular biology of insect sodium channels and pyrethroid resistance
- PMID: 24704279
- PMCID: PMC4484874
- DOI: 10.1016/j.ibmb.2014年03月01日2
Molecular biology of insect sodium channels and pyrethroid resistance
Abstract
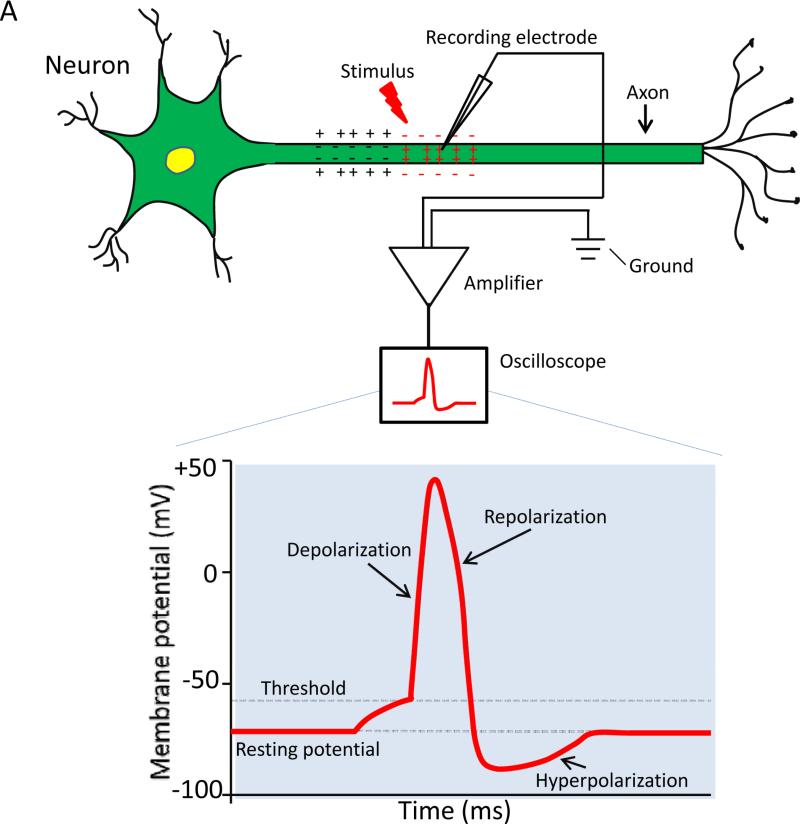
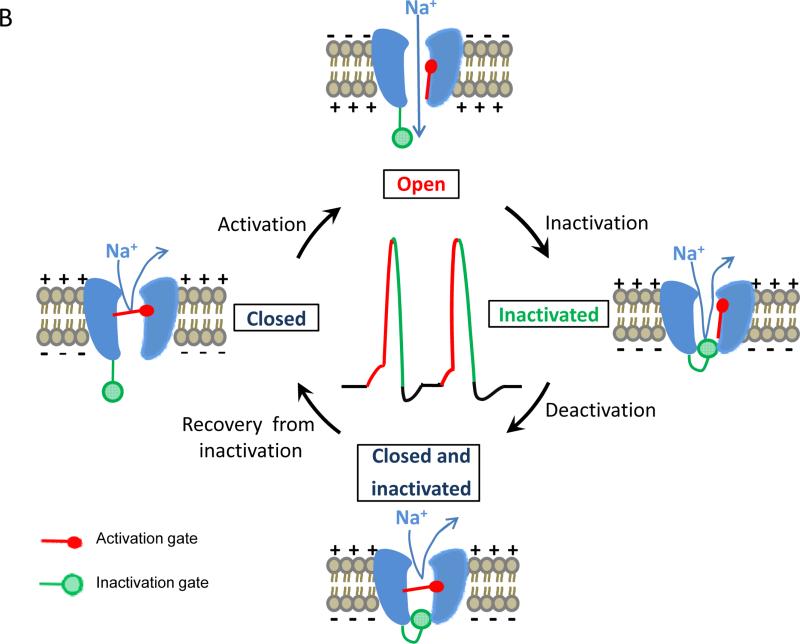
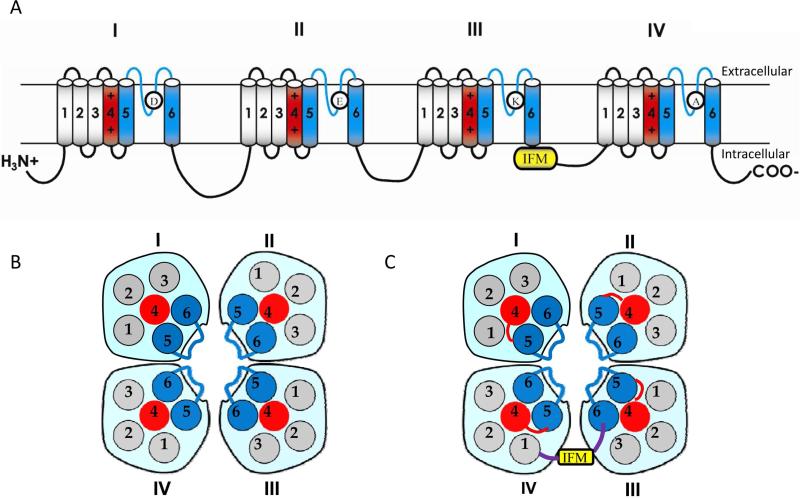
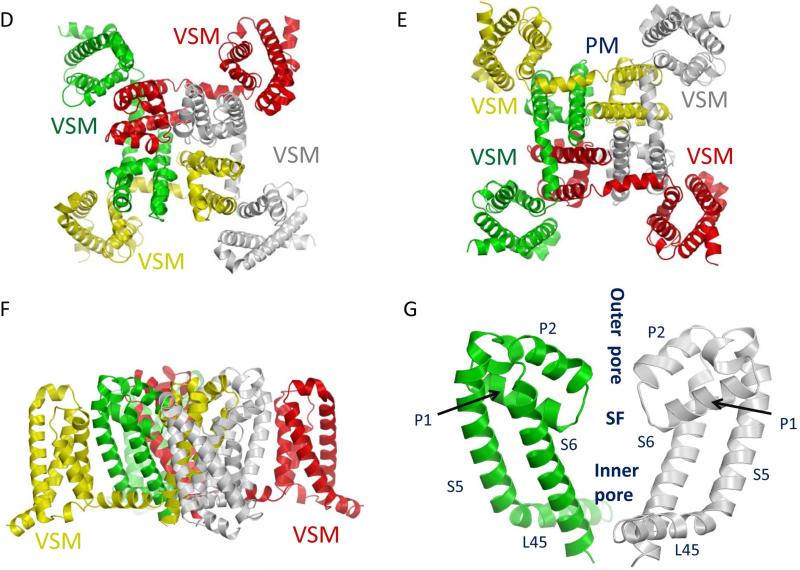
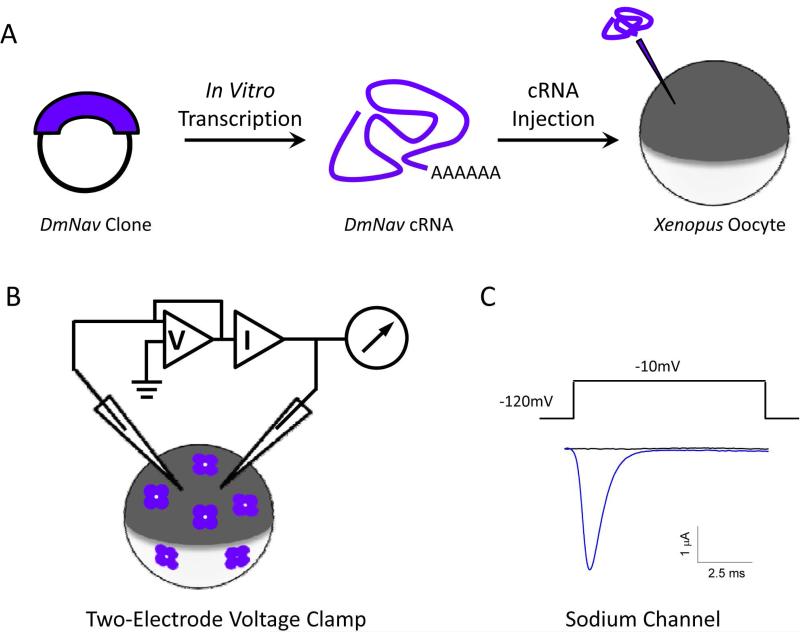
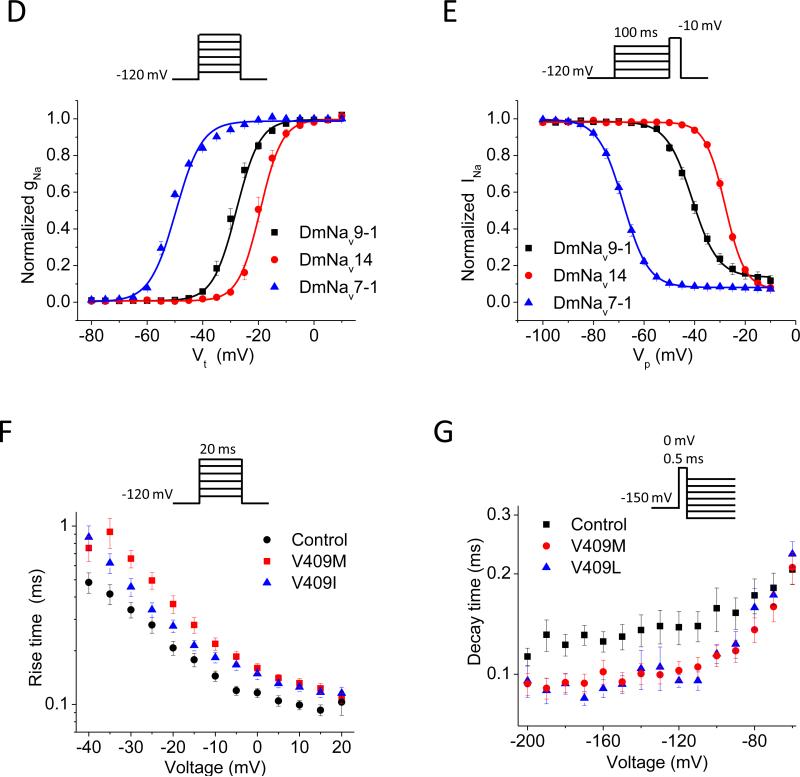
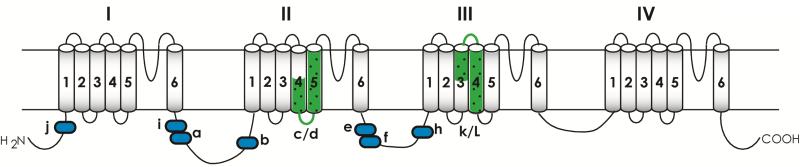
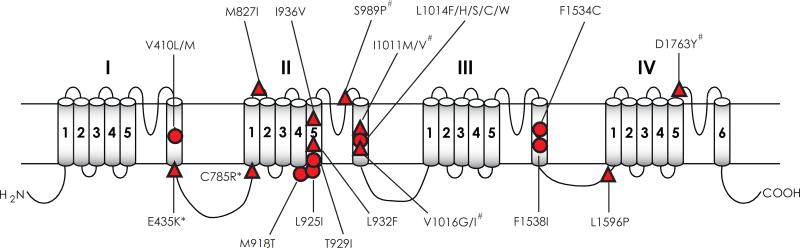
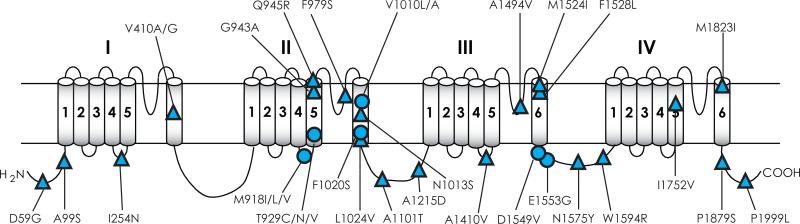
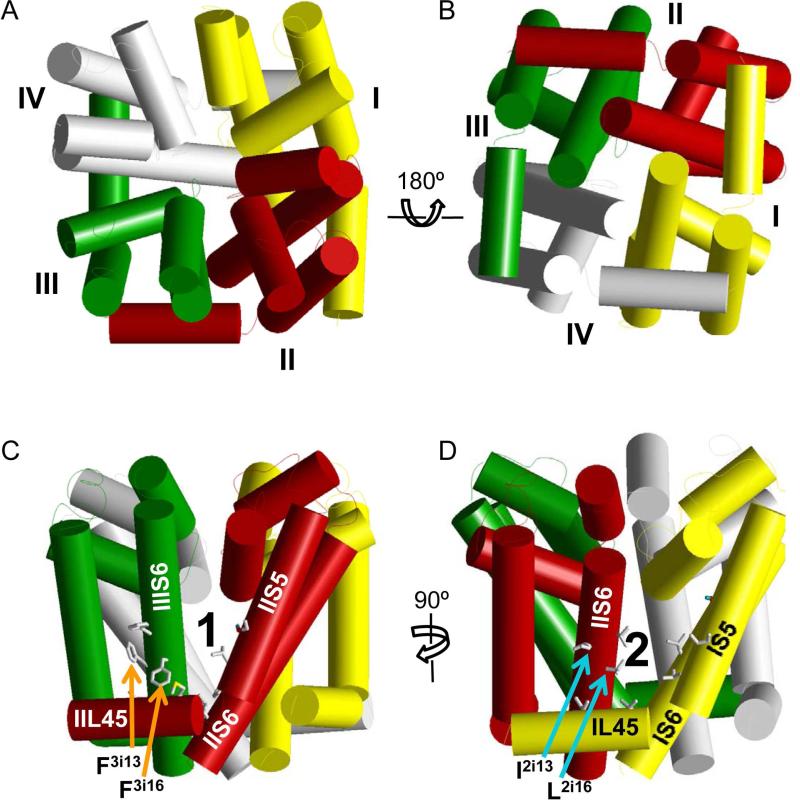
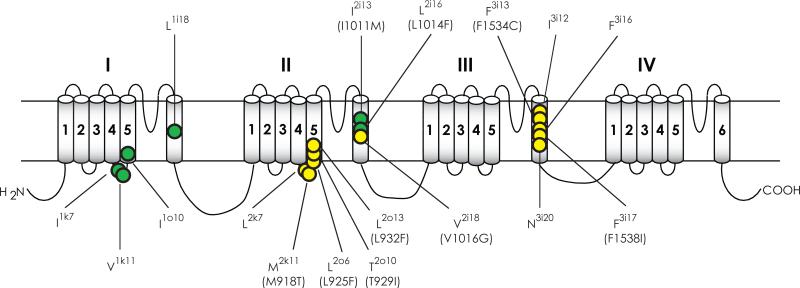
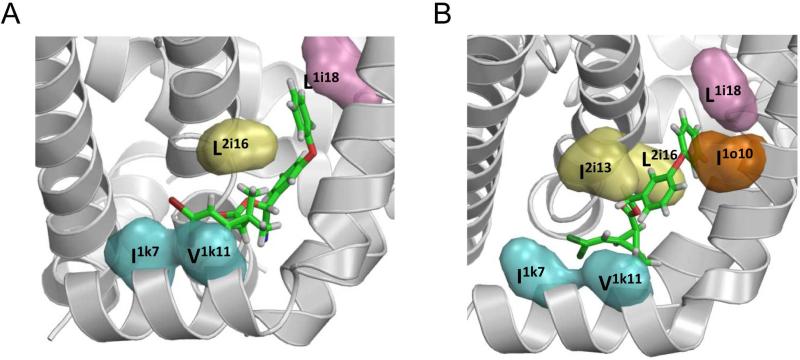
Voltage-gated sodium channels are essential for the initiation and propagation of the action potential in neurons and other excitable cells. Because of their critical roles in electrical signaling, sodium channels are targets of a variety of naturally occurring and synthetic neurotoxins, including several classes of insecticides. This review is intended to provide an update on the molecular biology of insect sodium channels and the molecular mechanism of pyrethroid resistance. Although mammalian and insect sodium channels share fundamental topological and functional properties, most insect species carry only one sodium channel gene, compared to multiple sodium channel genes found in each mammalian species. Recent studies showed that two posttranscriptional mechanisms, alternative splicing and RNA editing, are involved in generating functional diversity of sodium channels in insects. More than 50 sodium channel mutations have been identified to be responsible for or associated with knockdown resistance (kdr) to pyrethroids in various arthropod pests and disease vectors. Elucidation of molecular mechanism of kdr led to the identification of dual receptor sites of pyrethroids on insect sodium channels. Many of the kdr mutations appear to be located within or close to the two receptor sites. The accumulating knowledge of insect sodium channels and their interactions with insecticides provides a foundation for understanding the neurophysiology of sodium channels in vivo and the development of new and safer insecticides for effective control of arthropod pests and human disease vectors.
Keywords: Alternative splicing; Knockdown resistance; Pyrethroid receptor sites; Pyrethroids; RNA editing; Sodium channel.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Bass BL. RNA editing. Oxford University Press; 2001.
-
- Bourdin CM, Moignot B, Wang L, Murillo L, Juchaux M, Quinchard S, Lapied B, Guerineau NC, Dong K, Legros C. Intron retention in mRNA encoding ancillary subunit of insect voltage-gated sodium channel modulates channel expression, gating regulation and drug sensitivity. PLoS One. 2013;8:e67290. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources