Invasive non-typhoidal Salmonella typhimurium ST313 are not host-restricted and have an invasive phenotype in experimentally infected chickens
- PMID: 24130915
- PMCID: PMC3794976
- DOI: 10.1371/journal.pntd.0002487
Invasive non-typhoidal Salmonella typhimurium ST313 are not host-restricted and have an invasive phenotype in experimentally infected chickens
Abstract
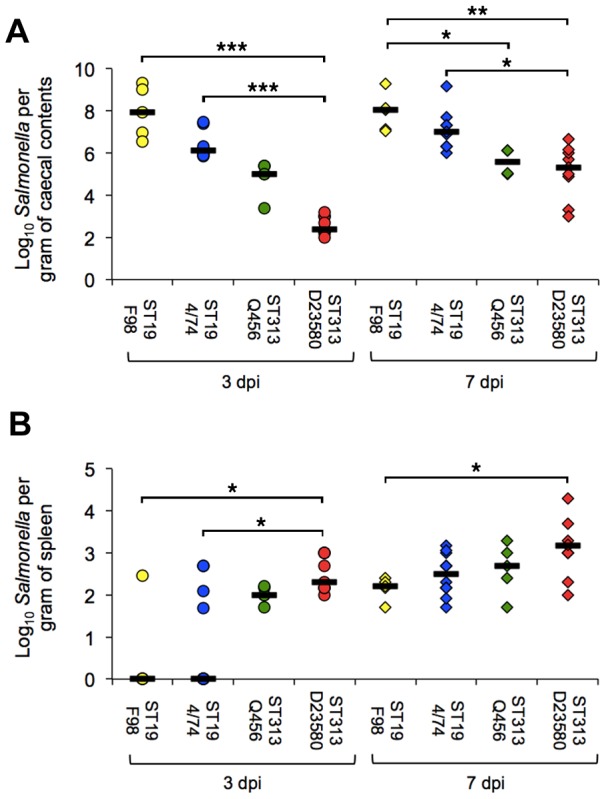
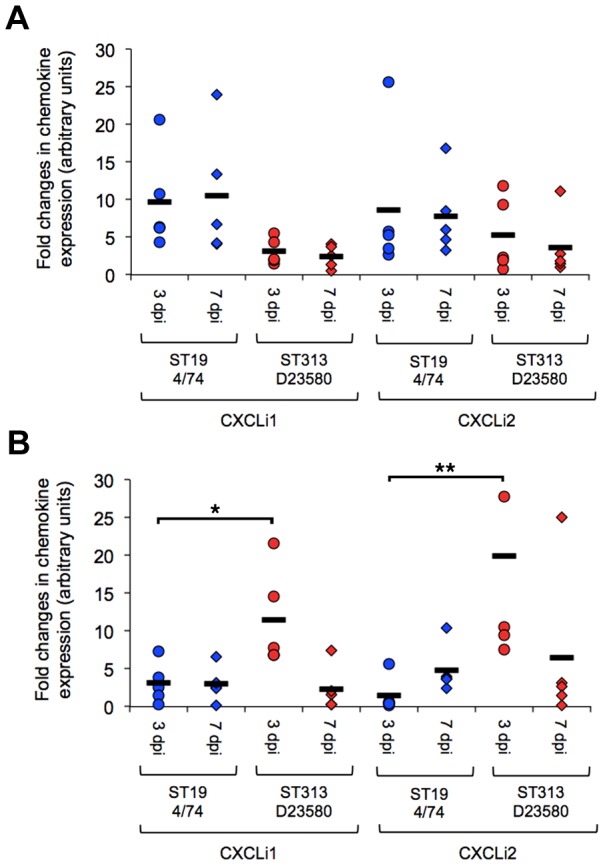
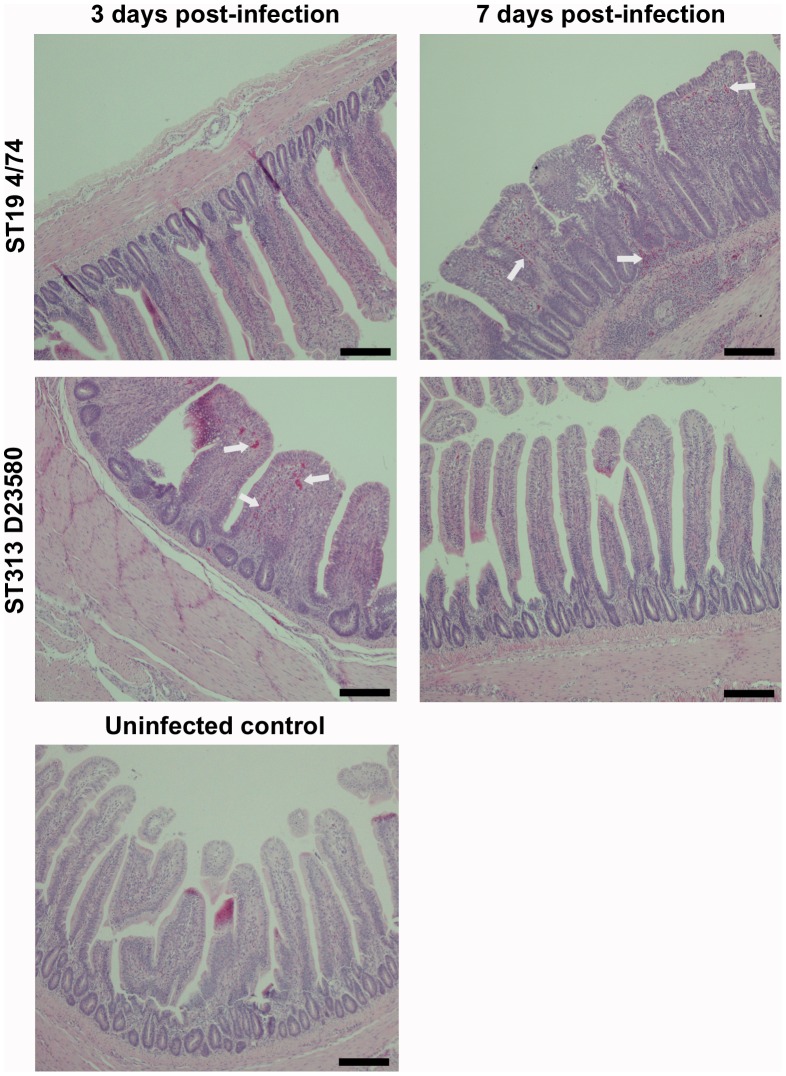
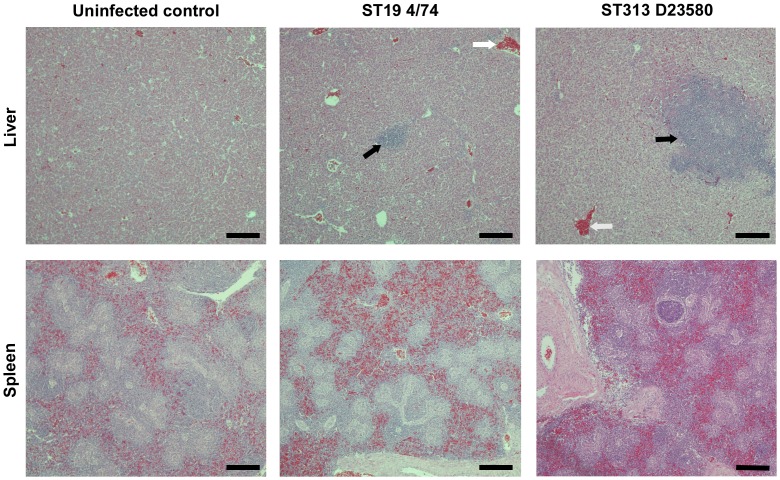
Salmonella enterica serovar Typhimurium Sequence Type (ST) 313 is a major cause of invasive non-Typhoidal salmonellosis in sub-Saharan Africa. No animal reservoir has been identified, and it has been suggested that ST313 is adapted to humans and transmission may occur via person-to-person spread. Here, we show that ST313 cause severe invasive infection in chickens as well as humans. Oral infection of chickens with ST313 isolates D23580 and Q456 resulted in rapid infection of spleen and liver with all birds infected at these sites by 3 days post-infection. In contrast, the well-defined ST19 S. Typhimurium isolates F98 and 4/74 were slower to cause invasive disease. Both ST19 and ST313 caused hepatosplenomegaly, and this was most pronounced in the ST313-infected animals. At 3 and 7 days post-infection, colonization of the gastrointestinal tract was lower in birds infected with the ST313 isolates compared with ST19. Histological examination and expression of CXCL chemokines in the ileum showed that both D23580 (ST313) and 4/74 (ST19) strains caused increased CXCL expression at 3 days post-infection, and this was significantly higher in the ileum of D23580 vs 4/74 infected birds. At 7 days post-infection, reduced chemokine expression occurred in the ileum of the D23580 but not 4/74-infected birds. Histological analysis showed that D23580 infection resulted in rapid inflammation and pathology including villous flattening and fusion at 3 days post-infection, and subsequent resolution by 7 days. In contrast, 4/74 induced less inflammation and pathology at 3 days post-infection. The data presented demonstrate that ST313 is capable of causing invasive disease in a non-human host. The rapid invasive nature of infection in the chicken, coupled with lower gastrointestinal colonization, supports the hypothesis that ST313 is a distinct pathovariant of S. Typhimurium that has evolved to become a systemic pathogen that can cause disease in several hosts.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical