Identification of a type IV secretion substrate of Brucella abortus that participates in the early stages of intracellular survival
- PMID: 24119283
- PMCID: PMC3945426
- DOI: 10.1111/cmi.12224
Identification of a type IV secretion substrate of Brucella abortus that participates in the early stages of intracellular survival
Abstract
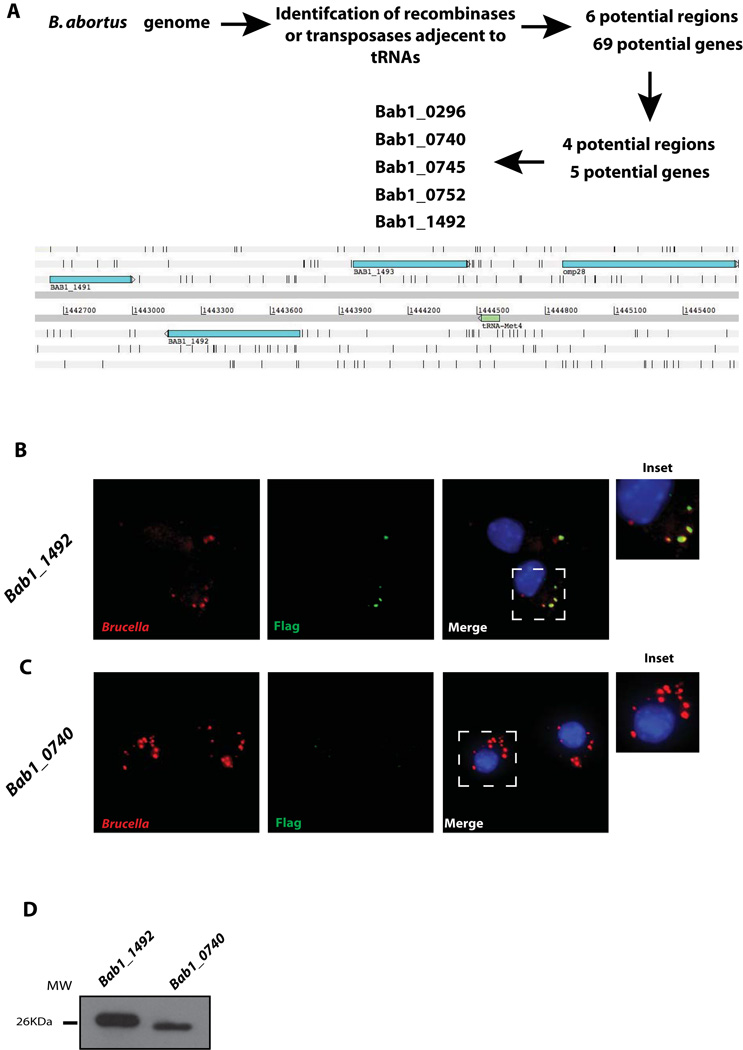
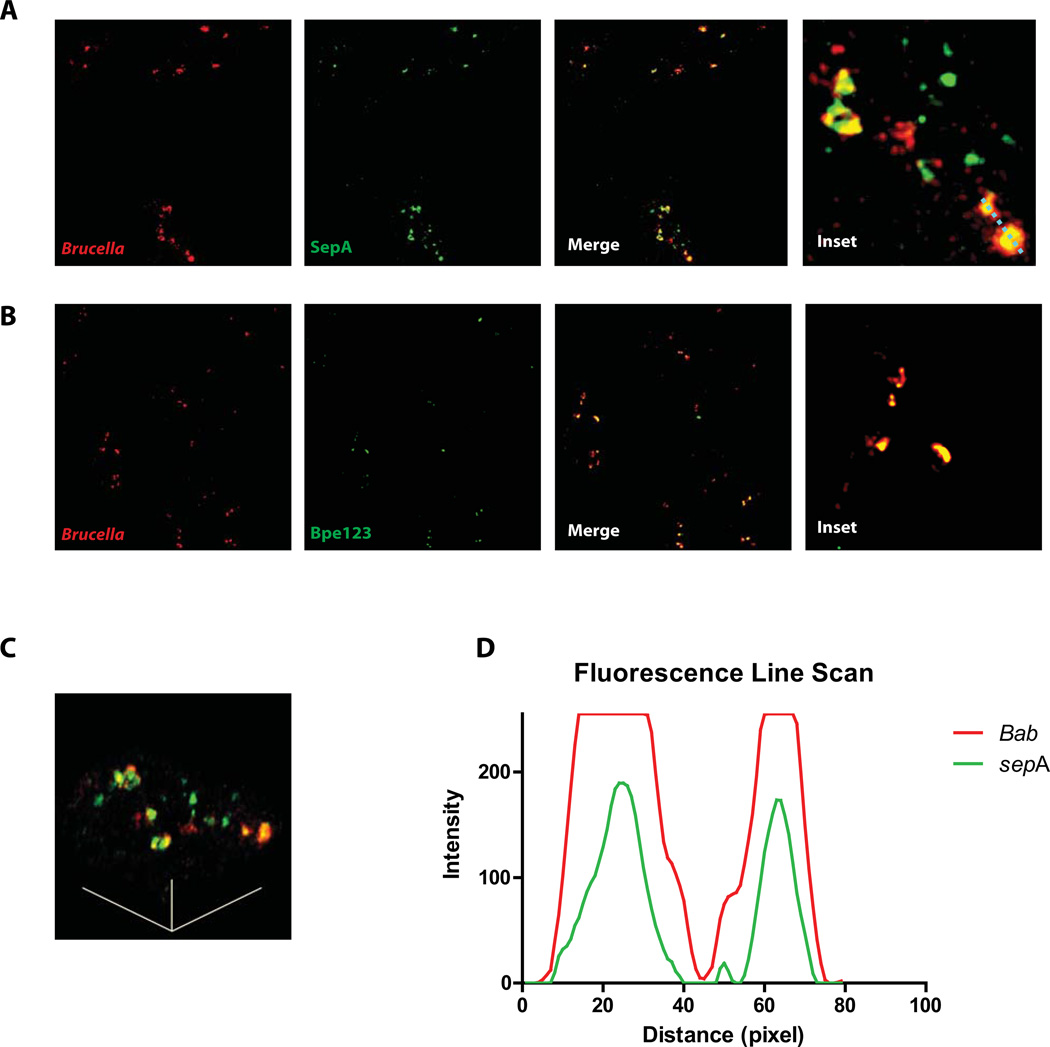
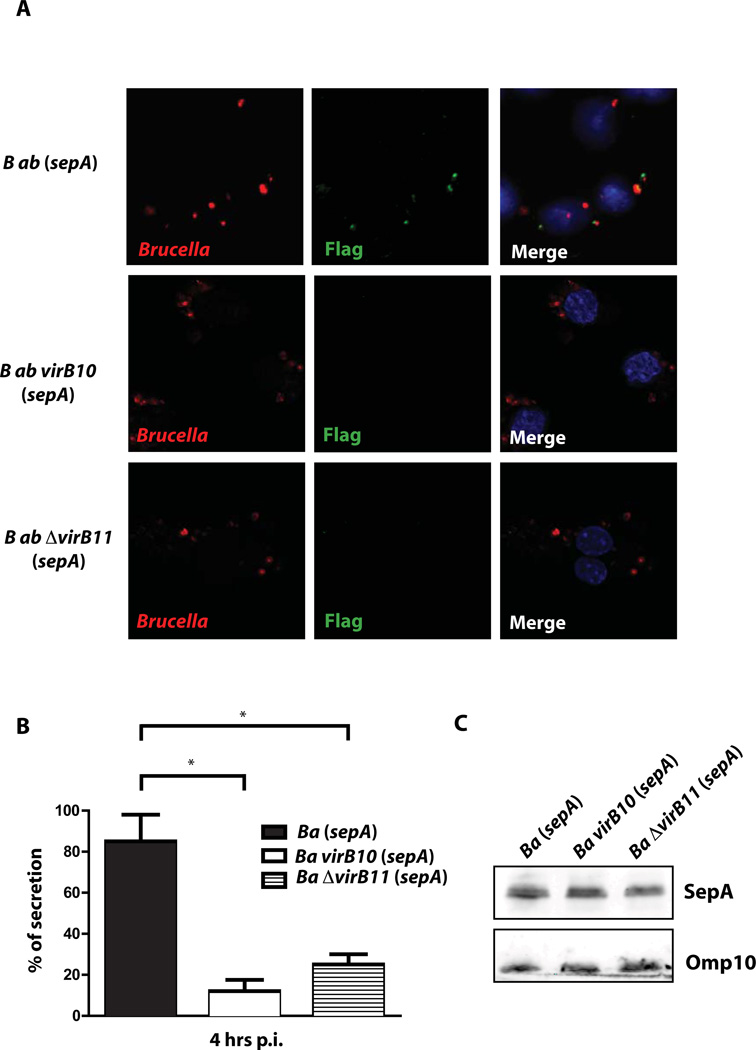
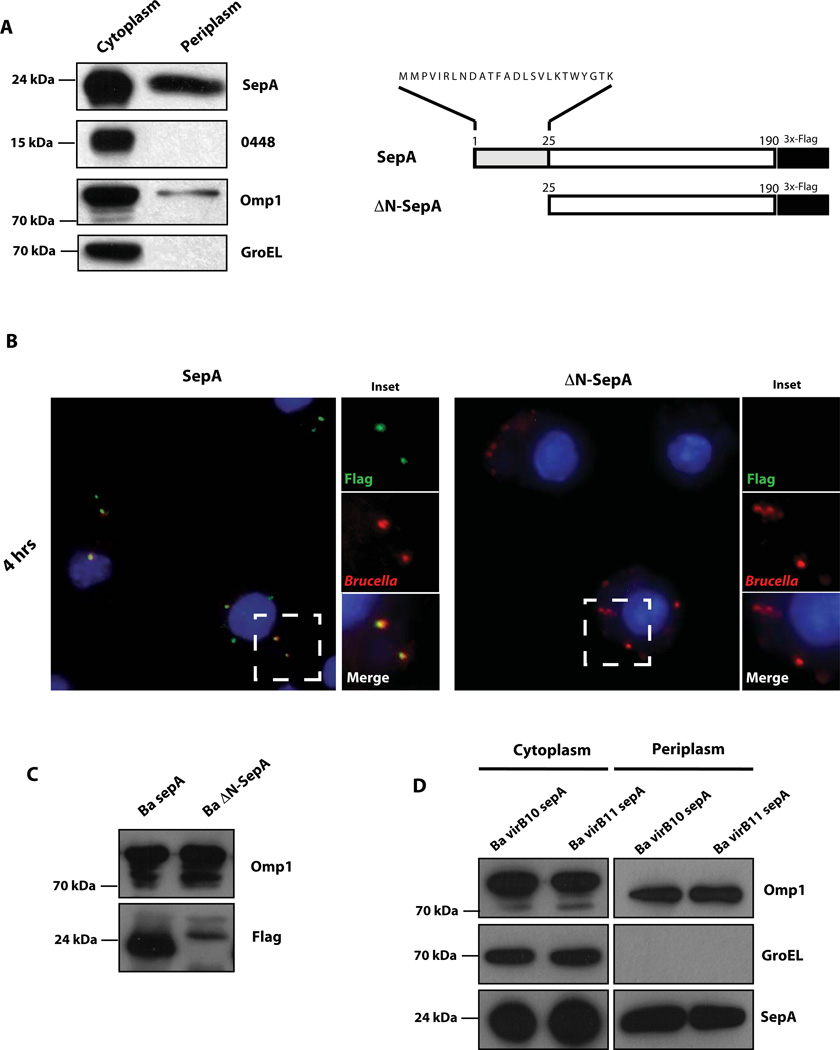
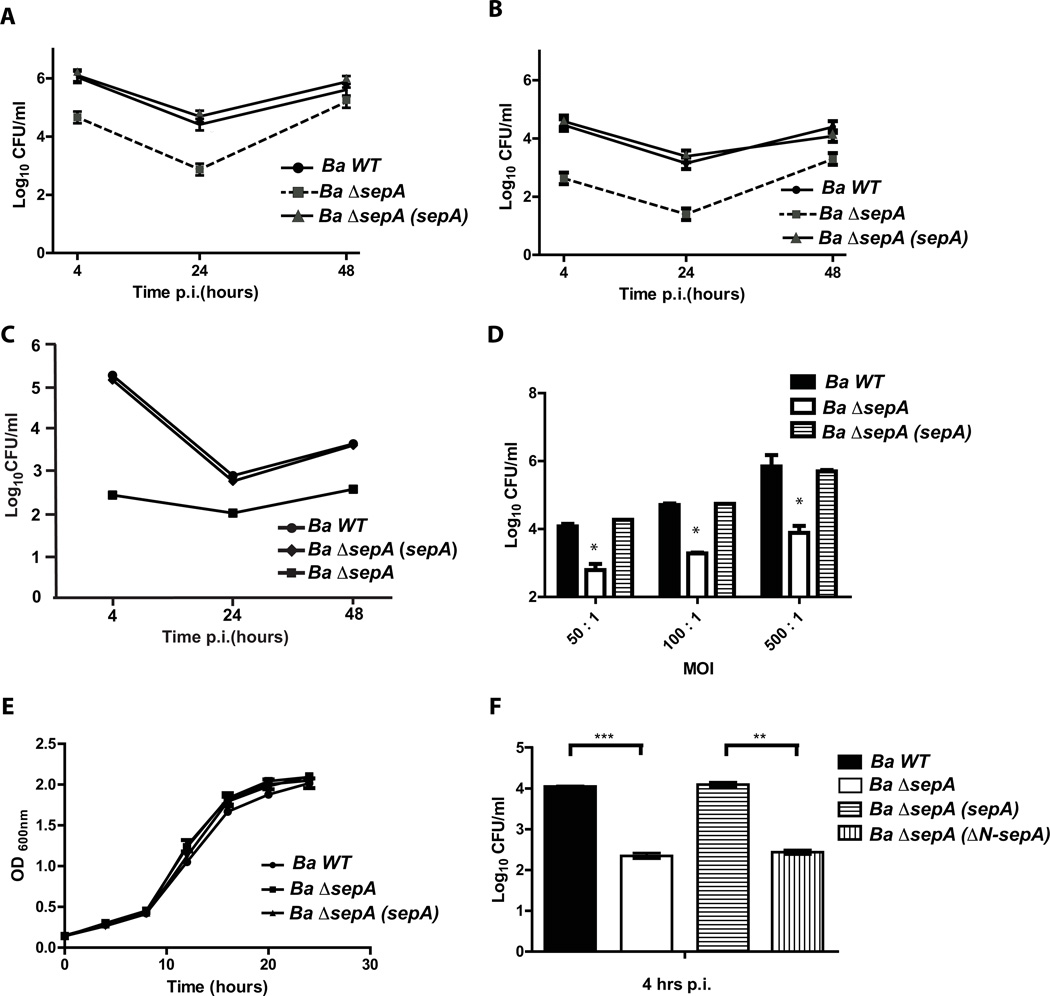
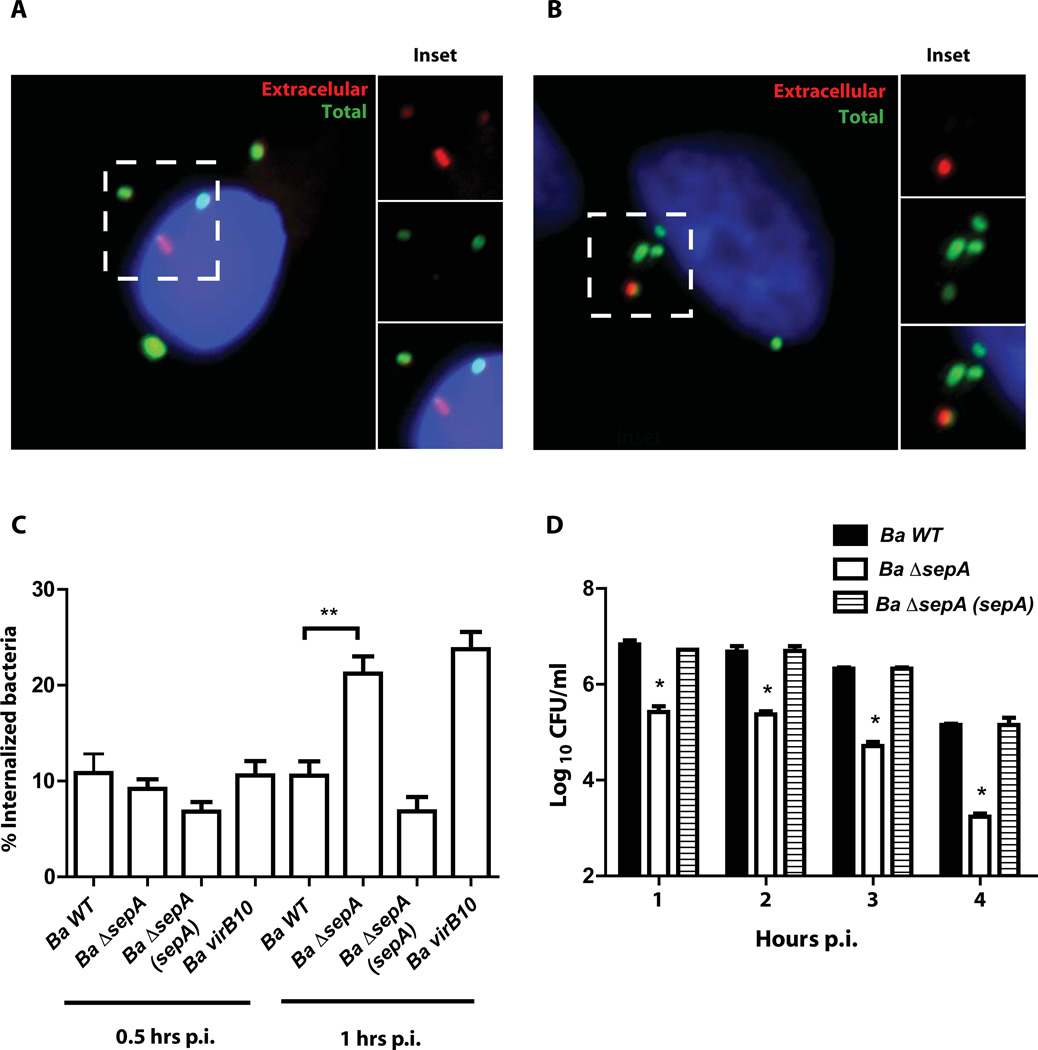
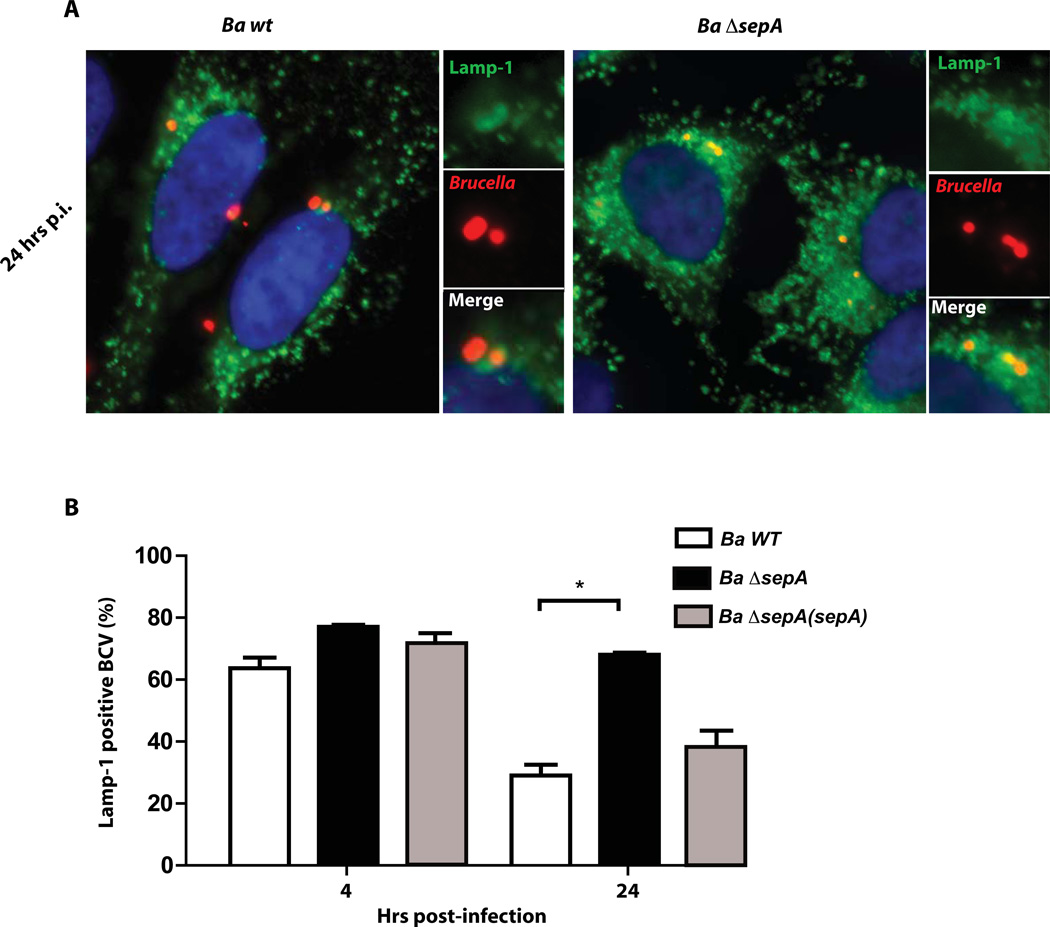
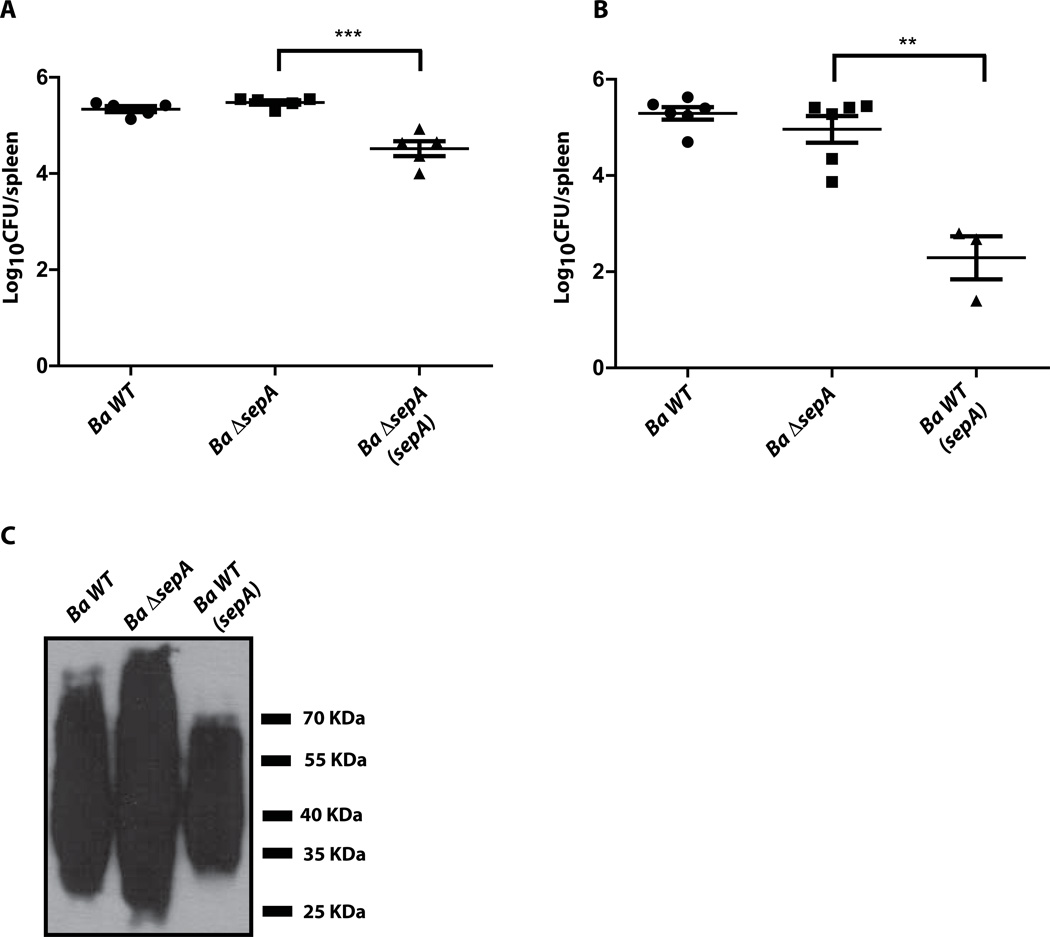
Brucella abortus, the aetiological agent of bovine brucellosis, is an intracellular pathogen whose virulence is completely dependent on a type IV secretion system. This secretion system translocates effector proteins into the host cell to modulate the intracellular fate of the bacterium in order to establish a secure niche were it actively replicates. Although much has been done in understanding how this secretion system participates in the virulence process, few effector proteins have been identified to date. We describe here the identification of a type IV secretion substrate (SepA) that is only present in Brucella spp. and has no detectable homology to known proteins. This protein is secreted in a virB-dependent manner in a two-step process involving a periplasmic intermediate and secretion is necessary for its function. The deletion mutant showed a defect in the early stages of intracellular replication in professional and non-professional phagocytes although it invades the cells more efficiently than the wild-type parental strain. Our results indicate that, even though the mutant was more invasive, it had a defect in excluding the lysosomal marker Lamp-1 and was inactivated more efficiently during the early phases of the intracellular life cycle.
© 2013 John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous