The dauer hypothesis and the evolution of parasitism: 20 years on and still going strong
- PMID: 24095839
- PMCID: PMC3947200
- DOI: 10.1016/j.ijpara.201308004
The dauer hypothesis and the evolution of parasitism: 20 years on and still going strong
Abstract
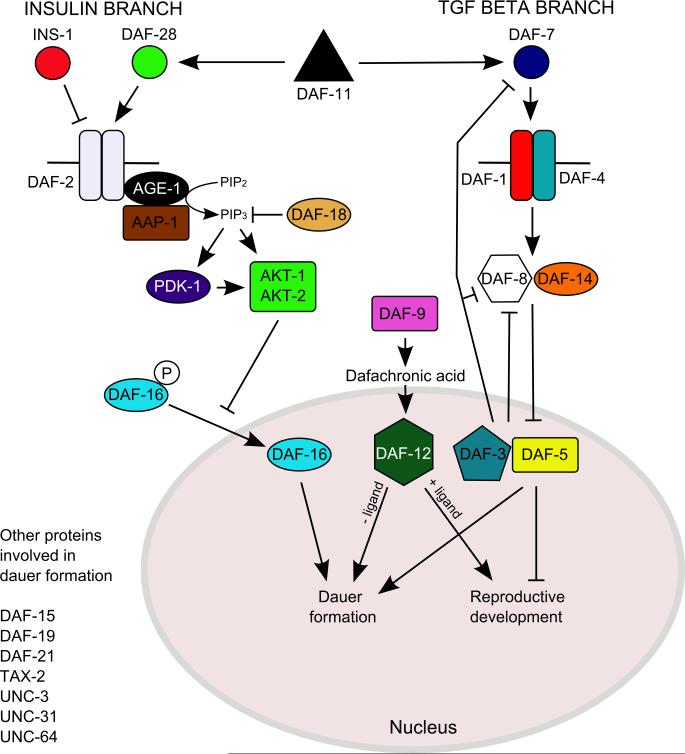
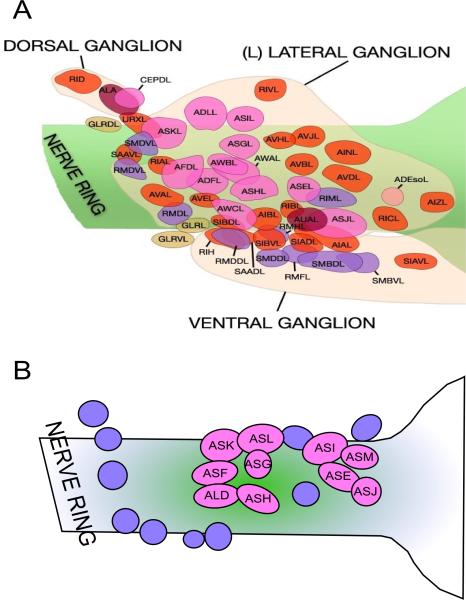
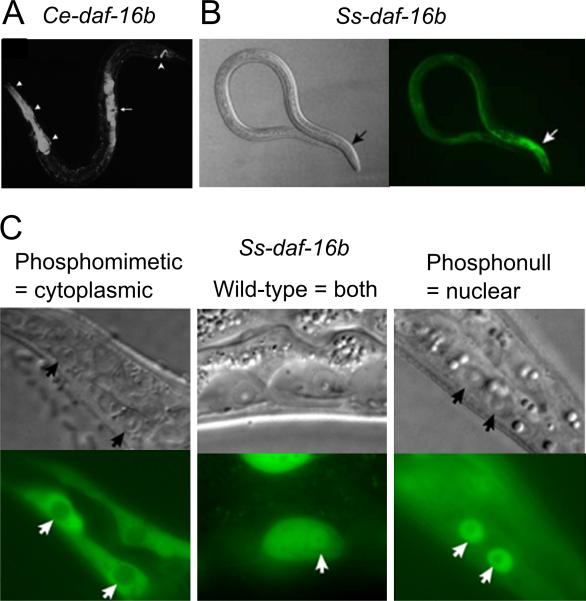
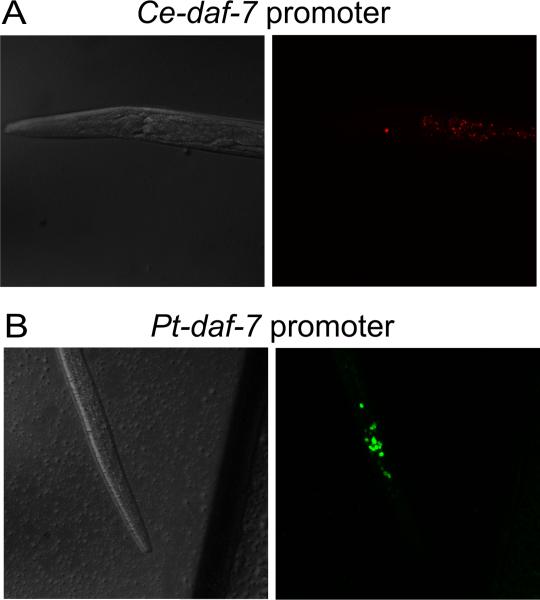
How any complex trait has evolved is a fascinating question, yet the evolution of parasitism among the nematodes is arguably one of the most arresting. How did free-living nematodes cross that seemingly insurmountable evolutionary chasm between soil dwelling and survival inside another organism? Which of the many finely honed responses to the varied and harsh environments of free-living nematodes provided the material upon which natural selection could act? Although several complementary theories explain this phenomenon, I will focus on the dauer hypothesis. The dauer hypothesis posits that the arrested third-stage dauer larvae of free-living nematodes such as Caenorhabditis elegans are, due to their many physiological similarities with infective third-stage larvae of parasitic nematodes, a pre-adaptation to parasitism. If so, then a logical extension of this hypothesis is that the molecular pathways which control entry into and recovery from dauer formation by free-living nematodes in response to environmental cues have been co-opted to control the processes of infective larval arrest and activation in parasitic nematodes. The molecular machinery that controls dauer entry and exit is present in a wide range of parasitic nematodes. However, the developmental outputs of the different pathways are both conserved and divergent, not only between populations of C. elegans or between C. elegans and parasitic nematodes but also between different species of parasitic nematodes. Thus the picture that emerges is more nuanced than originally predicted and may provide insights into the evolution of such an interesting and complex trait.
Keywords: Co-option; Dauer hypothesis; Evolution; Insulin signalling; TGF-β.
Copyright © 2013 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Altun ZF, Hall DH. Handbook of C. elegans anatomy.In: Wormatlas. 2008 http://www.wormatlas.org/hermaphrodite/hermaphroditehomepage.htm.
-
- Ashton FT, Bhopale VM, Fine AE, Schad GA. Sensory neuroanatomy of a skin-penetrating nematode parasite: Strongyloides stercoralis. I. Amphidial neurons. J Comp Neurol. 1995;357:281–295. - PubMed
-
- Ashton FT, Bhopale VM, Holt D, Smith G, Schad GA. Developmental switching in the parasitic nematode Strongyloides stercoralis is controlled by the ASF and ASI amphidial neurons. J Parasitol. 1998;84:691–695. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources