Precision mapping of the human O-GalNAc glycoproteome through SimpleCell technology
- PMID: 23584533
- PMCID: PMC3655468
- DOI: 10.1038/emboj.2013.79
Precision mapping of the human O-GalNAc glycoproteome through SimpleCell technology
Abstract
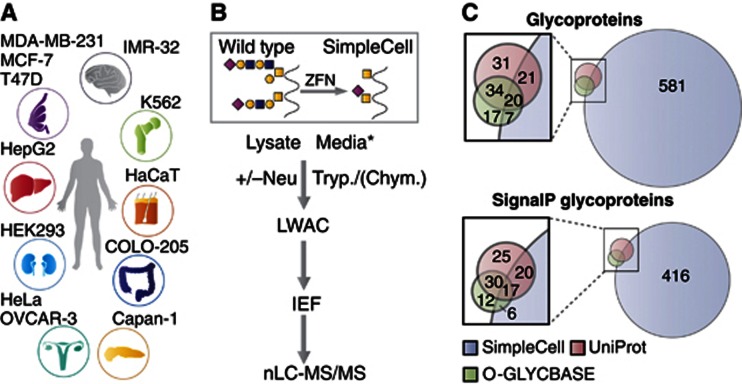
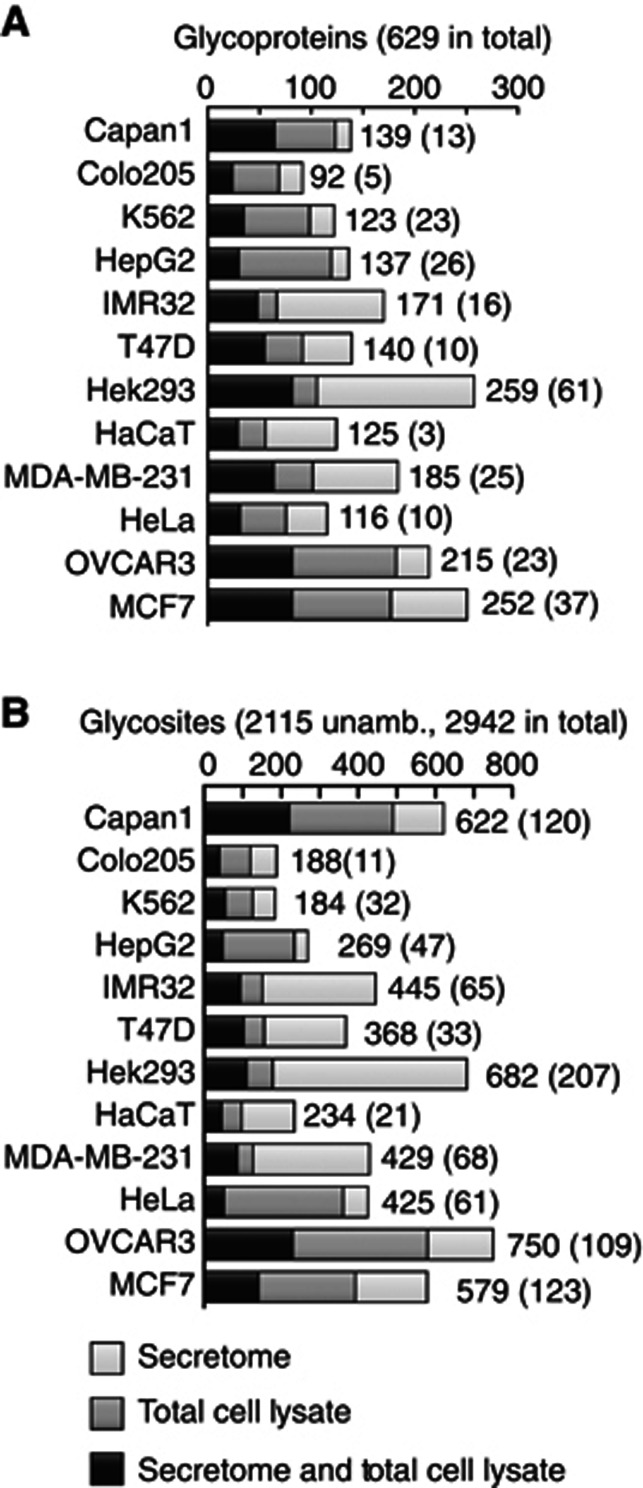
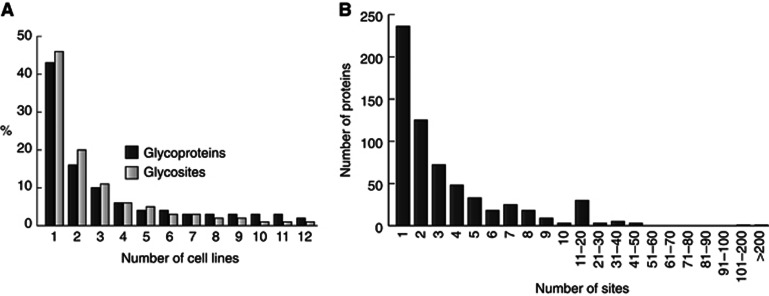
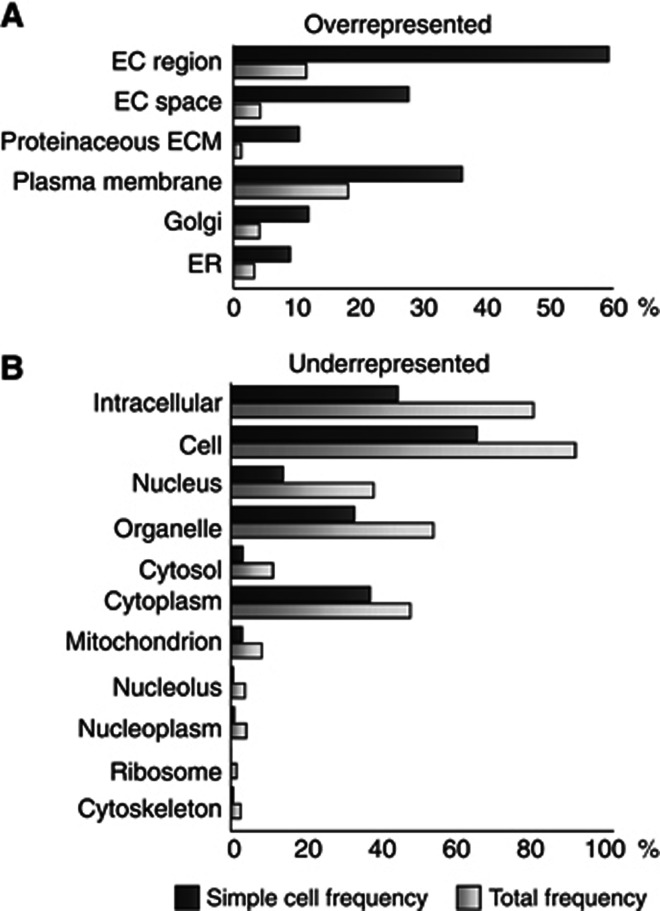
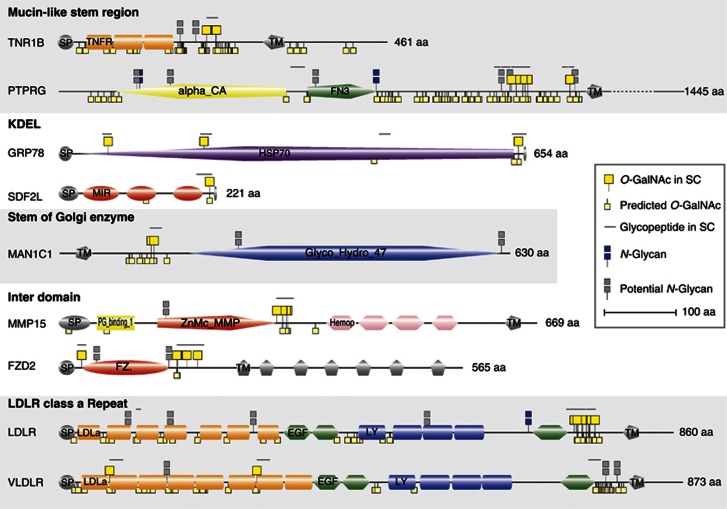
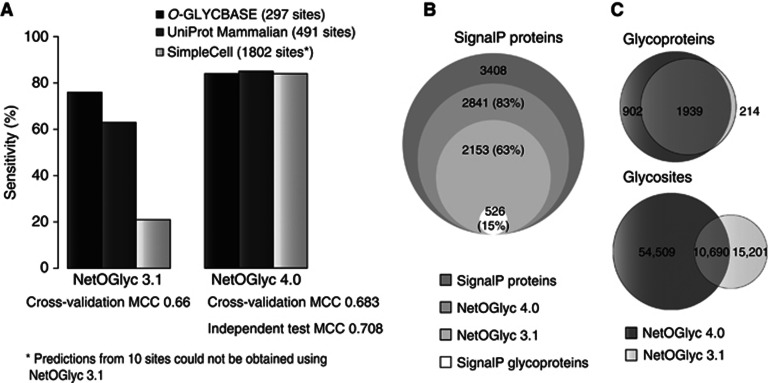
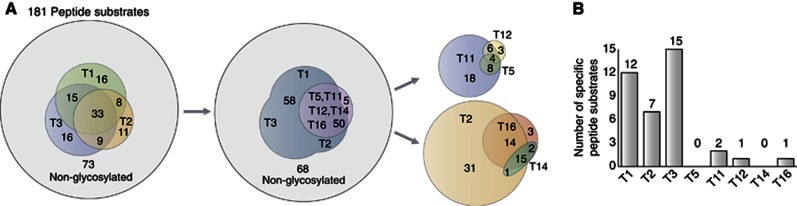
Glycosylation is the most abundant and diverse posttranslational modification of proteins. While several types of glycosylation can be predicted by the protein sequence context, and substantial knowledge of these glycoproteomes is available, our knowledge of the GalNAc-type O-glycosylation is highly limited. This type of glycosylation is unique in being regulated by 20 polypeptide GalNAc-transferases attaching the initiating GalNAc monosaccharides to Ser and Thr (and likely some Tyr) residues. We have developed a genetic engineering approach using human cell lines to simplify O-glycosylation (SimpleCells) that enables proteome-wide discovery of O-glycan sites using 'bottom-up' ETD-based mass spectrometric analysis. We implemented this on 12 human cell lines from different organs, and present a first map of the human O-glycoproteome with almost 3000 glycosites in over 600 O-glycoproteins as well as an improved NetOGlyc4.0 model for prediction of O-glycosylation. The finding of unique subsets of O-glycoproteins in each cell line provides evidence that the O-glycoproteome is differentially regulated and dynamic. The greatly expanded view of the O-glycoproteome should facilitate the exploration of how site-specific O-glycosylation regulates protein function.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Alfaro JF, Gong CX, Monroe ME, Aldrich JT, Clauss TR, Purvine SO, Wang Z, Camp DG 2nd, Shabanowitz J, Stanley P, Hart GW, Hunt DF, Yang F, Smith RD (2012) Tandem mass spectrometry identifies many mouse brain O-GlcNAcylated proteins including EGF domain-specific O-GlcNAc transferase targets. Proc Natl Acad Sci USA 109: 7280–7285 - PMC - PubMed
-
- Bennett EP, Hassan H, Clausen H (1996) cDNA cloning and expression of a novel human UDP-N-acetyl-alpha-D-galactosamine polypeptide N-acetylgalactosaminyltransferase, GalNAc-T3. J Biol Chem 271: 17006–17012 - PubMed
-
- Davis CG, Elhammer A, Russell DW, Schneider WJ, Kornfeld S, Brown MS, Goldstein JL (1986) Deletion of clustered O-linked carbohydrates does not impair function of low density lipoprotein receptor in transfected fibroblasts. J Biol Chem 261: 2828–2838 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials