Aberrant assembly of RNA recognition motif 1 links to pathogenic conversion of TAR DNA-binding protein of 43 kDa (TDP-43)
- PMID: 23558684
- PMCID: PMC3663511
- DOI: 10.1074/jbc.M113.451849
Aberrant assembly of RNA recognition motif 1 links to pathogenic conversion of TAR DNA-binding protein of 43 kDa (TDP-43)
Abstract
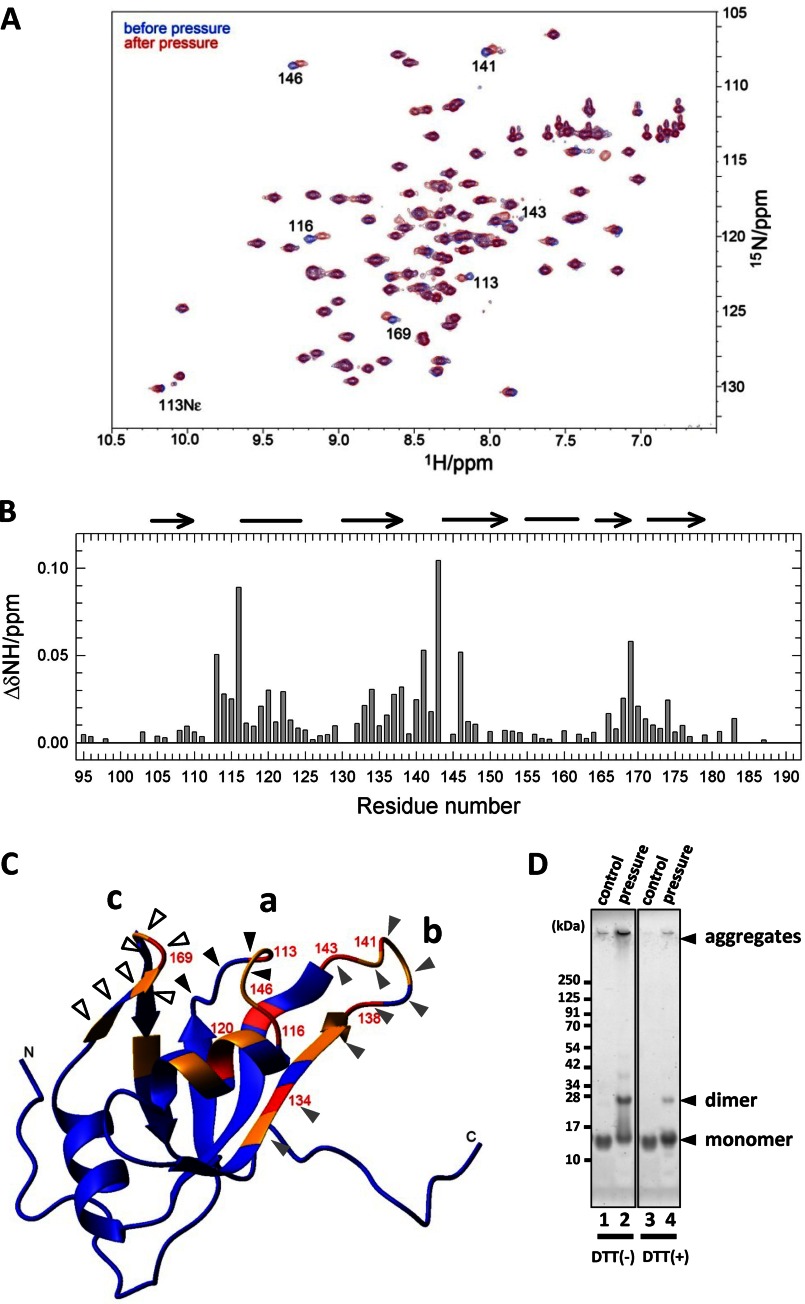
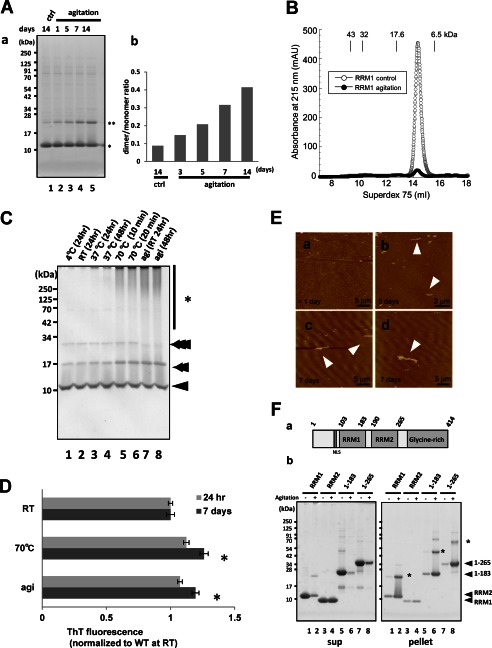
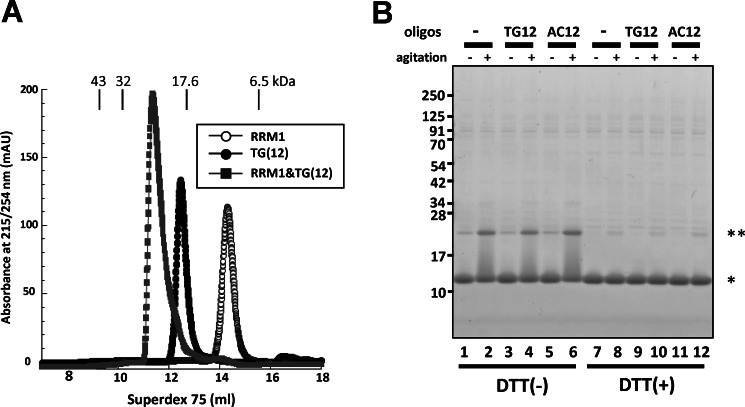
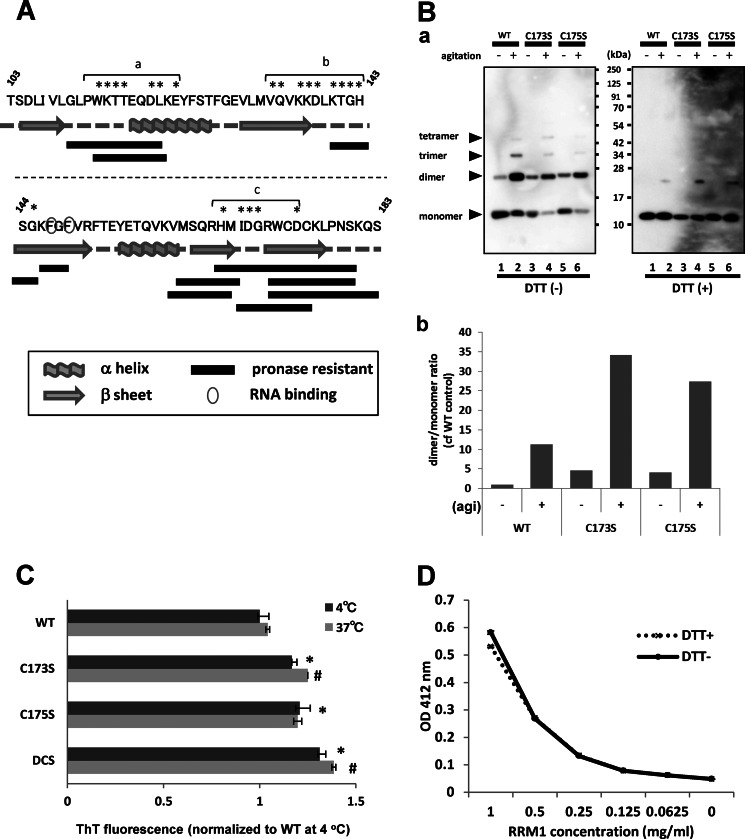
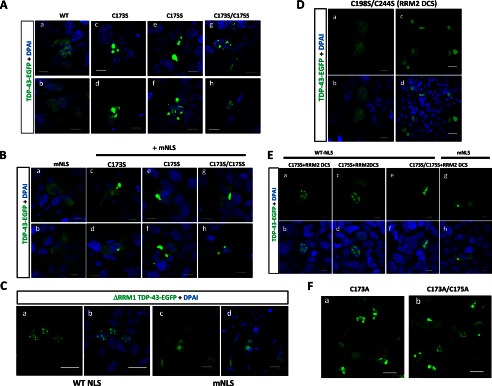
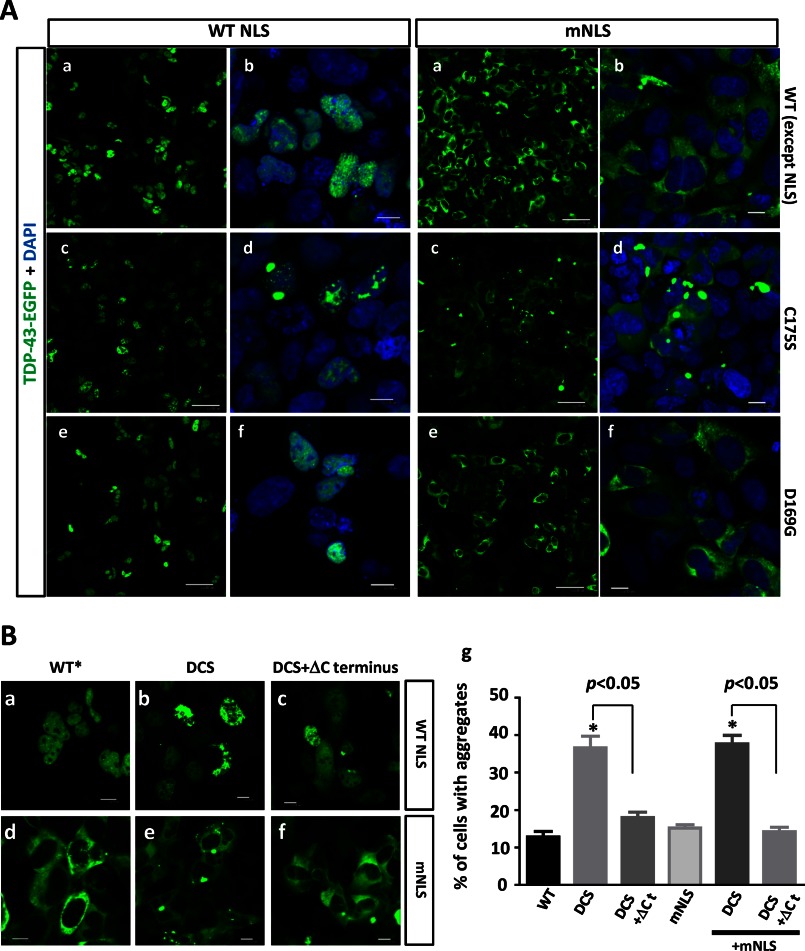
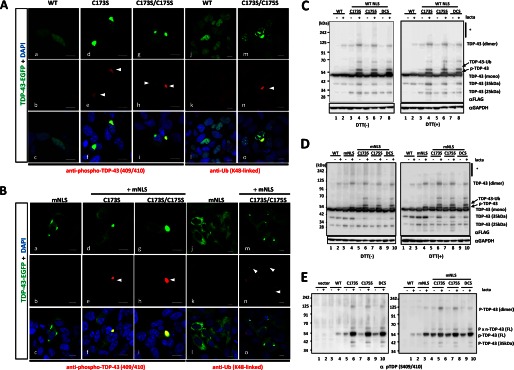
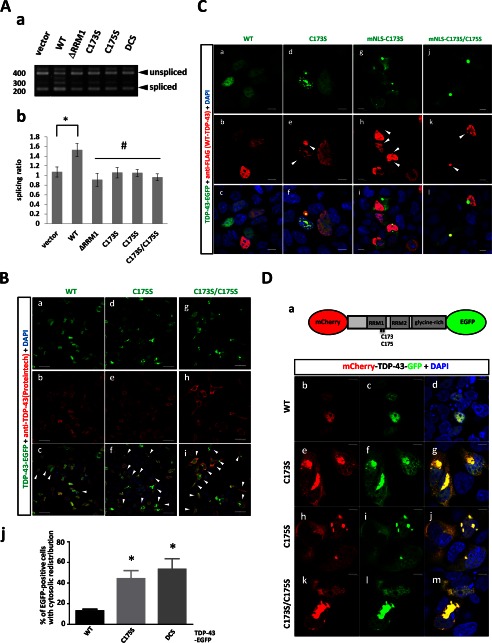
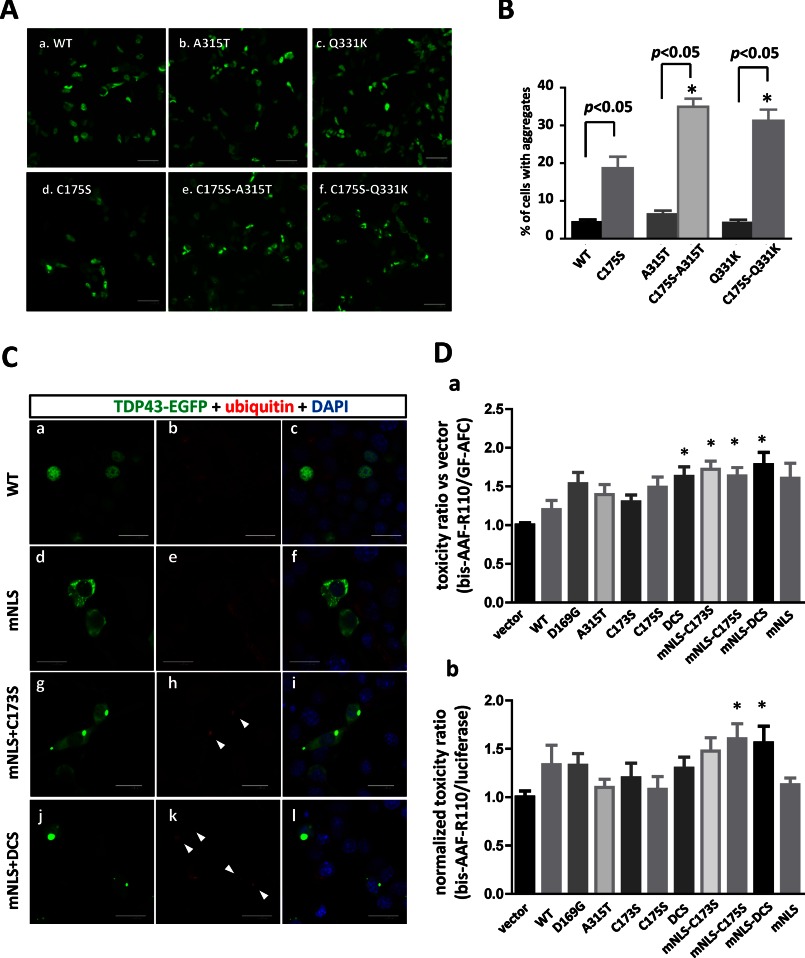
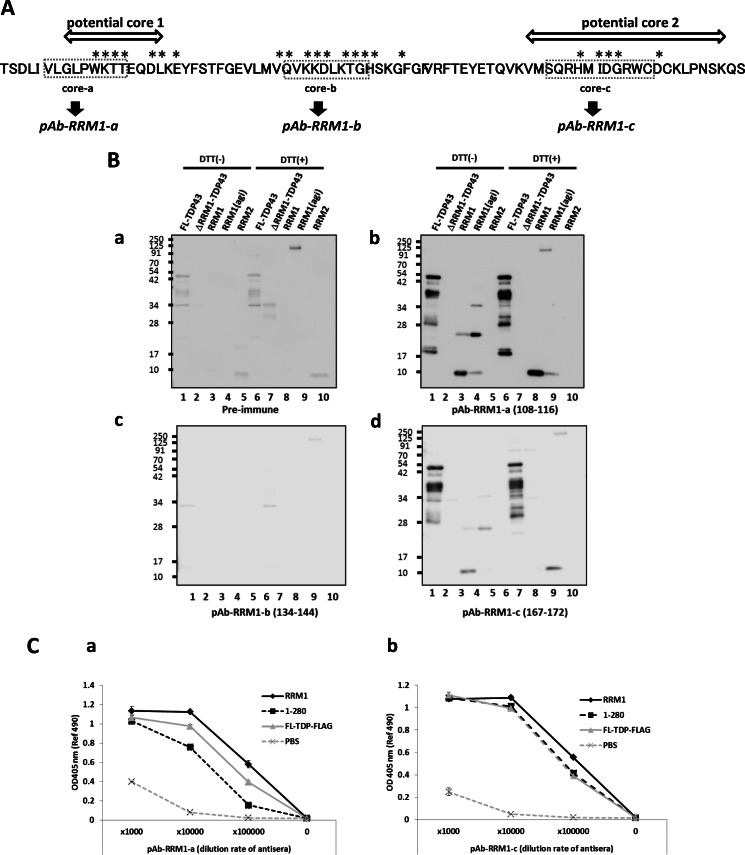
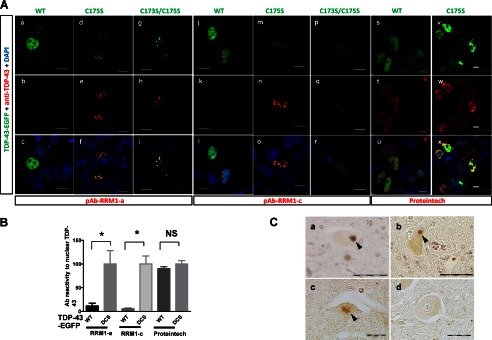
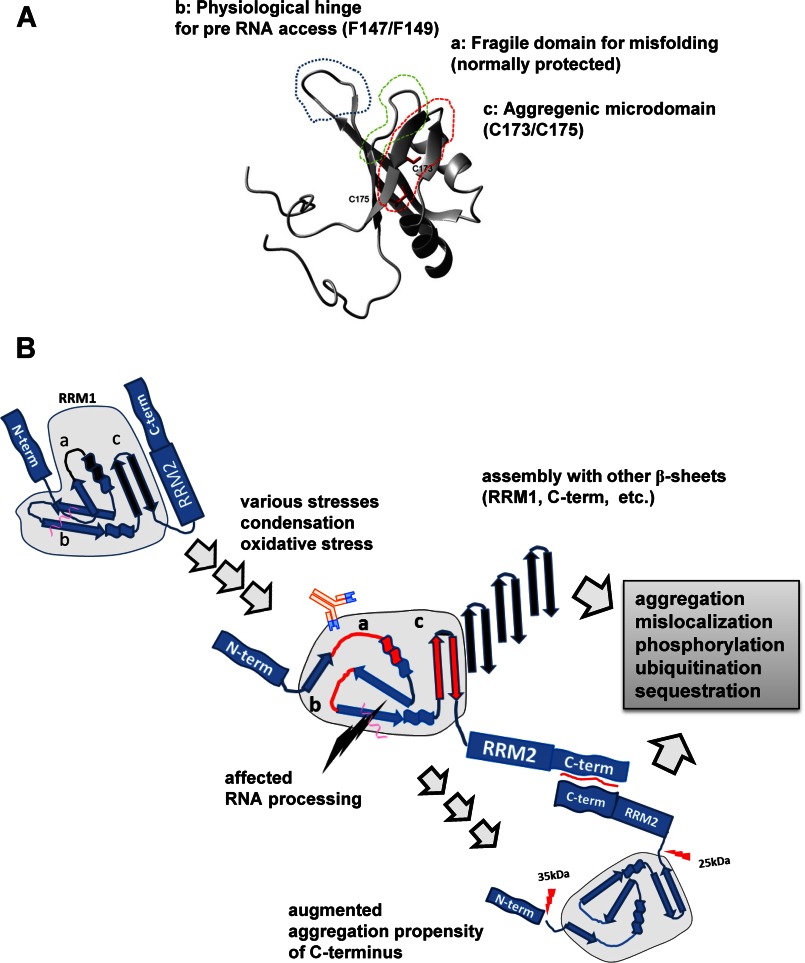
Aggregation of TAR DNA-binding protein of 43 kDa (TDP-43) is a pathological signature of amyotrophic lateral sclerosis (ALS). Although accumulating evidence suggests the involvement of RNA recognition motifs (RRMs) in TDP-43 proteinopathy, it remains unclear how native TDP-43 is converted to pathogenic forms. To elucidate the role of homeostasis of RRM1 structure in ALS pathogenesis, conformations of RRM1 under high pressure were monitored by NMR. We first found that RRM1 was prone to aggregation and had three regions showing stable chemical shifts during misfolding. Moreover, mass spectrometric analysis of aggregated RRM1 revealed that one of the regions was located on protease-resistant β-strands containing two cysteines (Cys-173 and Cys-175), indicating that this region served as a core assembly interface in RRM1 aggregation. Although a fraction of RRM1 aggregates comprised disulfide-bonded oligomers, the substitution of cysteine(s) to serine(s) (C/S) resulted in unexpected acceleration of amyloid fibrils of RRM1 and disulfide-independent aggregate formation of full-length TDP-43. Notably, TDP-43 aggregates with RRM1-C/S required the C terminus, and replicated cytopathologies of ALS, including mislocalization, impaired RNA splicing, ubiquitination, phosphorylation, and motor neuron toxicity. Furthermore, RRM1-C/S accentuated inclusions of familial ALS-linked TDP-43 mutants in the C terminus. The relevance of RRM1-C/S-induced TDP-43 aggregates in ALS pathogenesis was verified by immunolabeling of inclusions of ALS patients and cultured cells overexpressing the RRM1-C/S TDP-43 with antibody targeting misfolding-relevant regions. Our results indicate that cysteines in RRM1 crucially govern the conformation of TDP-43, and aberrant self-assembly of RRM1 at amyloidogenic regions contributes to pathogenic conversion of TDP-43 in ALS.
Keywords: Amyotrophic Lateral Sclerosis (Lou Gehrig's Disease); Cell Biology; Protein Chemical Modification; Protein Misfolding; Structural Biology; TAR DNA-binding Protein 43 kDa.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Arai T., Hasegawa M., Akiyama H., Ikeda K., Nonaka T., Mori H., Mann D., Tsuchiya K., Yoshida M., Hashizume Y., Oda T. (2006) TDP-43 is a component of ubiquitin-positive tau-negative inclusions in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Biochem. Biophys. Res. Commun. 351, 602–611 - PubMed
-
- Neumann M., Sampathu D. M., Kwong L. K., Truax A. C., Micsenyi M. C., Chou T. T., Bruce J., Schuck T., Grossman M., Clark C. M., McCluskey L. F., Miller B. L., Masliah E., Mackenzie I. R., Feldman H., Feiden W., Kretzschmar H. A., Trojanowski J. Q., Lee V. M. (2006) Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science 314, 130–133 - PubMed
-
- Baloh R. H. (2012) How do the RNA-binding proteins TDP-43 and FUS relate to amyotrophic lateral sclerosis and frontotemporal degeneration, and to each other? Curr. Opin. Neurol. 25, 701–707 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous