Role of Cryptococcus neoformans Rho1 GTPases in the PKC1 signaling pathway in response to thermal stress
- PMID: 23159519
- PMCID: PMC3535842
- DOI: 10.1128/EC.05305-11
Role of Cryptococcus neoformans Rho1 GTPases in the PKC1 signaling pathway in response to thermal stress
Abstract
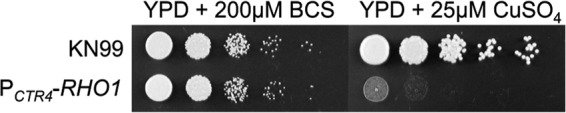
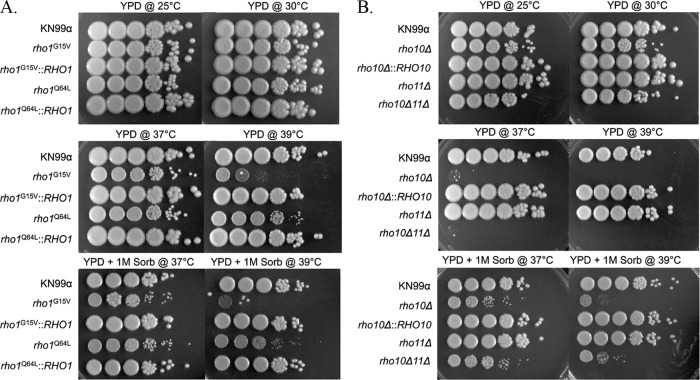
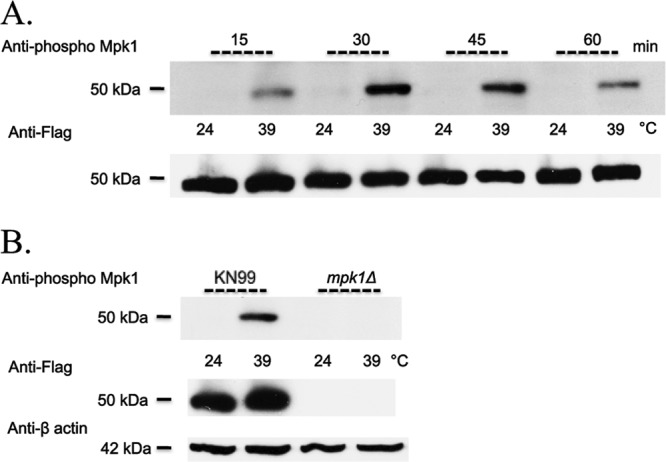
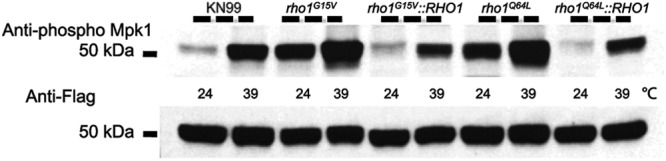
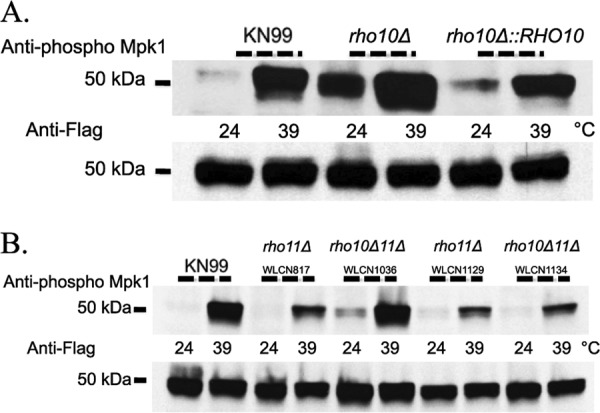
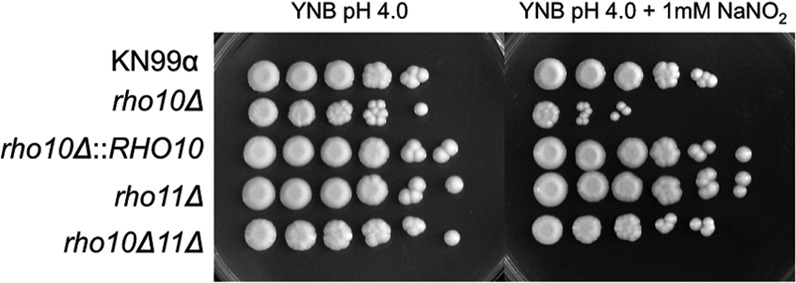
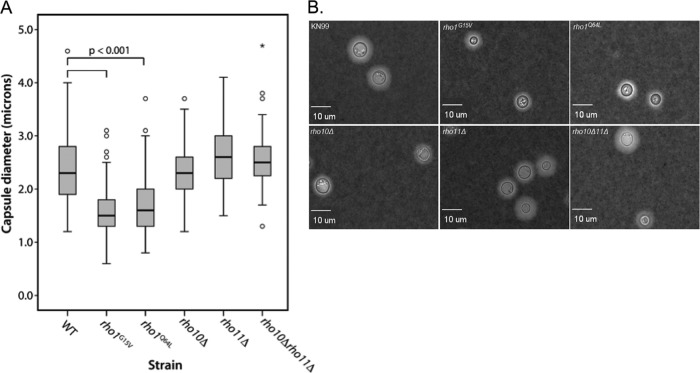
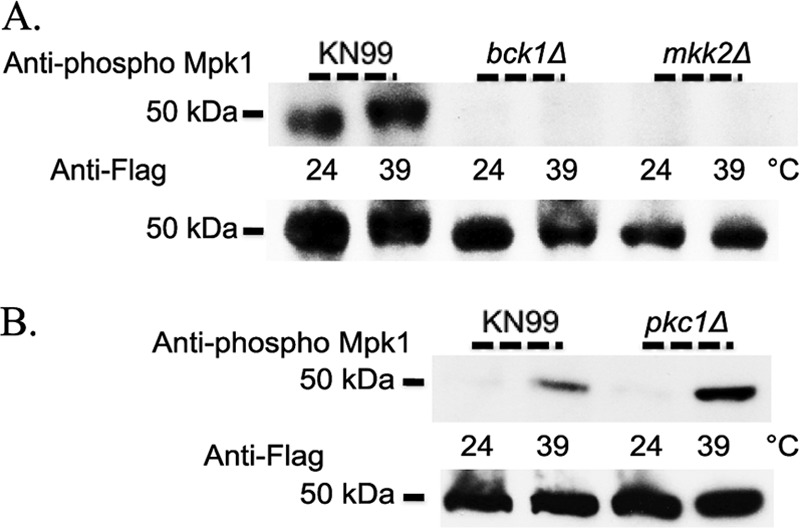
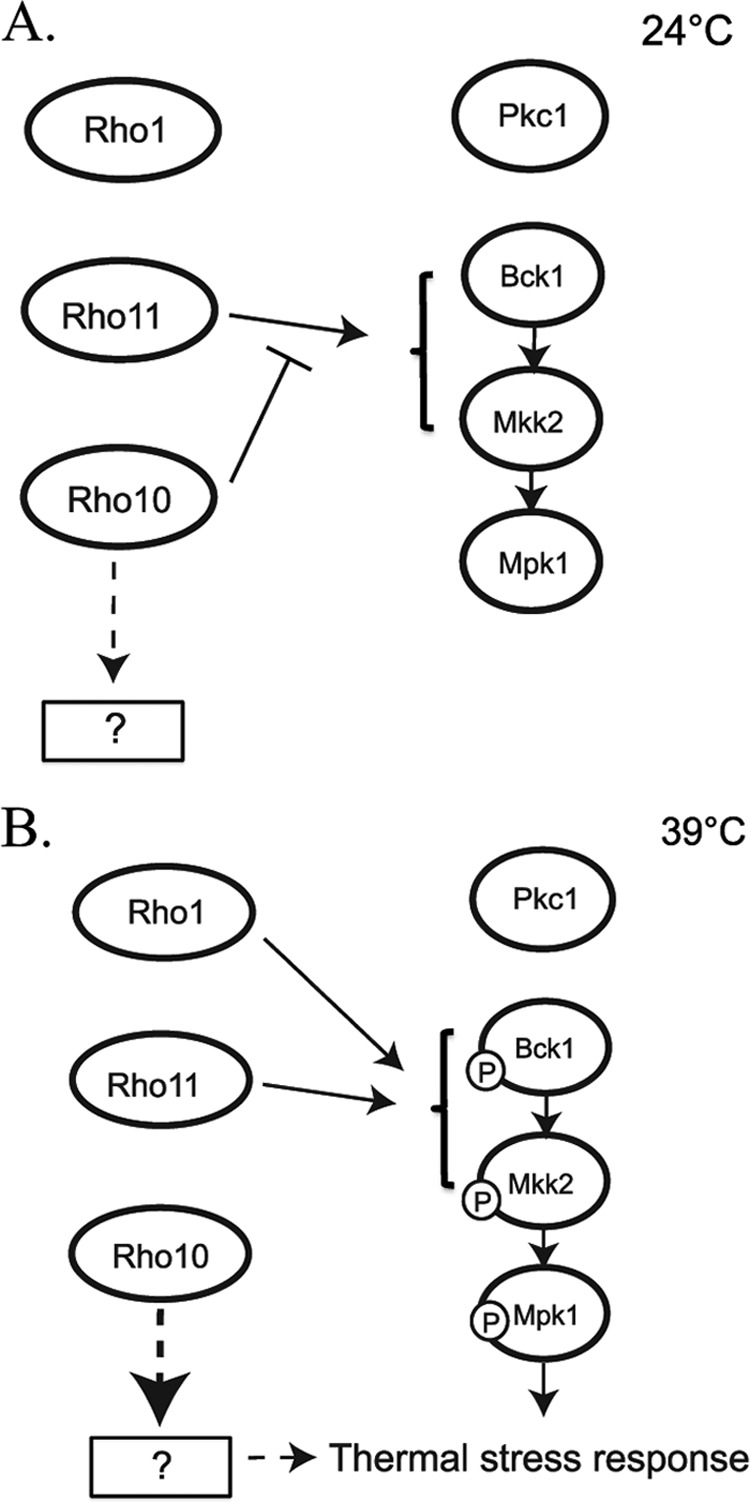
To initiate and establish infection in mammals, the opportunistic fungal pathogen Cryptococcus neoformans must survive and thrive upon subjection to host temperature. Primary maintenance of cell integrity is controlled through the protein kinase C1 (PKC1) signaling pathway, which is regulated by a Rho1 GTPase in Saccharomyces cerevisiae. We identified three C. neoformans Rho GTPases, Rho1, Rho10, and Rho11, and have begun to elucidate their role in growth and activation of the PKC1 pathway in response to thermal stress. Western blot analysis revealed that heat shock of wild-type cells resulted in phosphorylation of Mpk1 mitogen-activated protein kinase (MAPK). Constitutive activation of Rho1 caused phosphorylation of Mpk1 independent of temperature, indicating its role in pathway regulation. A strain with a deletion of RHO10 also displayed this constitutive Mpk1 phosphorylation phenotype, while one with a deletion of RHO11 yielded phosphorylation similar to that of wild type. Surprisingly, like a rho10Δ strain, a strain with a deletion of both RHO10 and RHO11 displayed temperature sensitivity but mimicked wild-type phosphorylation, which suggests that Rho10 and Rho11 have coordinately regulated functions. Heat shock-induced Mpk1 phosphorylation also required the PKC1 pathway kinases Bck1 and Mkk2. However, Pkc1, thought to be the major regulatory kinase of the cell integrity pathway, was dispensable for this response. Together, our results argue that Rho proteins likely interact via downstream components of the PKC1 pathway or by alternative pathways to activate the cell integrity pathway in C. neoformans.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Etienne-Manneville S, Hall A. 2002. Rho GTPases in cell biology. Nature 420:629–635 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous