Prolonged neutrophil dysfunction after Plasmodium falciparum malaria is related to hemolysis and heme oxygenase-1 induction
- PMID: 23100518
- PMCID: PMC3504608
- DOI: 10.4049/jimmunol.1201028
Prolonged neutrophil dysfunction after Plasmodium falciparum malaria is related to hemolysis and heme oxygenase-1 induction
Abstract
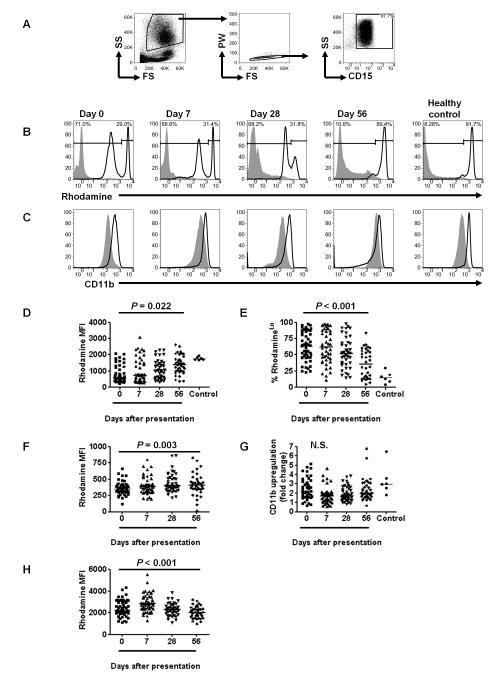
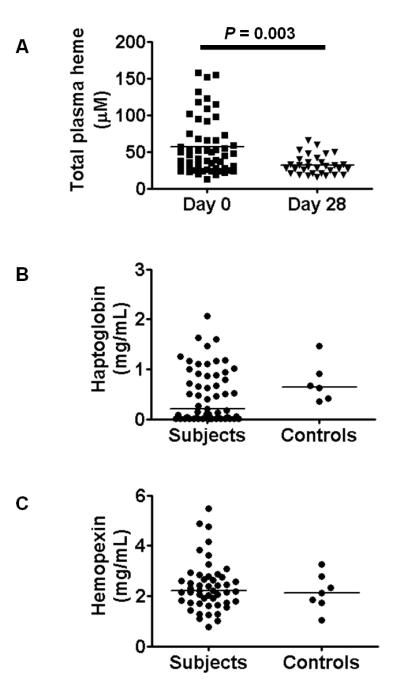
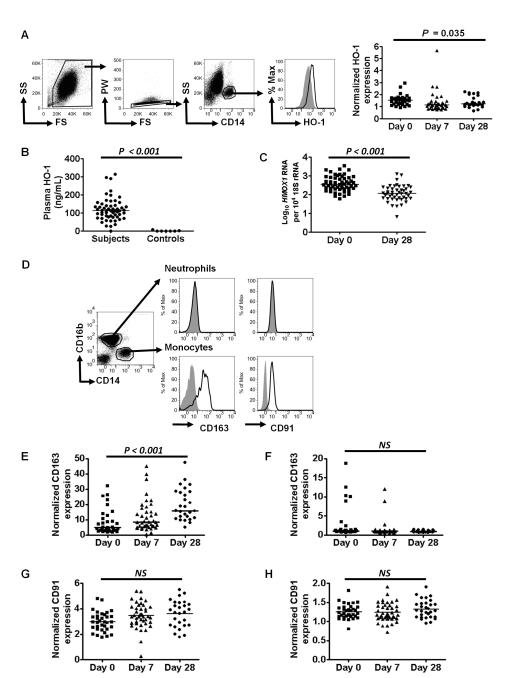
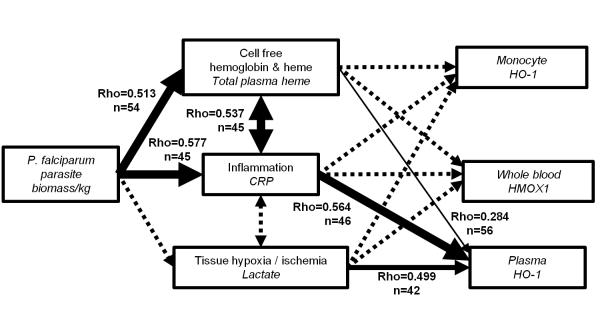
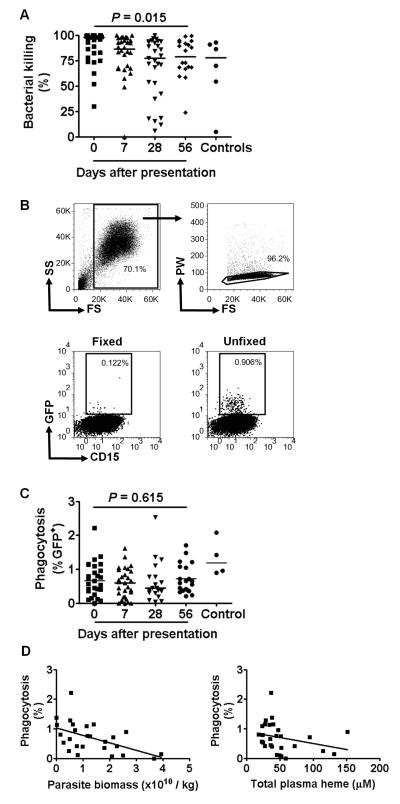
It is not known why people are more susceptible to bacterial infections such as nontyphoid Salmonella during and after a malaria infection, but in mice, malarial hemolysis impairs resistance to nontyphoid Salmonella by impairing the neutrophil oxidative burst. This acquired neutrophil dysfunction is a consequence of induction of the cytoprotective, heme-degrading enzyme heme oxygenase-1 (HO-1) in neutrophil progenitors in bone marrow. In this study, we assessed whether neutrophil dysfunction occurs in humans with malaria and how this relates to hemolysis. We evaluated neutrophil function in 58 Gambian children with Plasmodium falciparum malaria [55 (95%) with uncomplicated disease] and examined associations with erythrocyte count, haptoglobin, hemopexin, plasma heme, expression of receptors for heme uptake, and HO-1 induction. Malaria caused the appearance of a dominant population of neutrophils with reduced oxidative burst activity, which gradually normalized over 8 wk of follow-up. The degree of neutrophil impairment correlated significantly with markers of hemolysis and HO-1 induction. HO-1 expression was increased in blood during acute malaria, but at a cellular level HO-1 expression was modulated by changes in surface expression of the haptoglobin receptor (CD163). These findings demonstrate that neutrophil dysfunction occurs in P. falciparum malaria and support the relevance of the mechanistic studies in mice. Furthermore, they suggest the presence of a regulatory pathway to limit HO-1 induction by hemolysis in the context of infection and indicate new targets for therapeutic intervention to abrogate the susceptibility to bacterial infection in the context of hemolysis in humans.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- World malaria report 2011. World Health Organization; Geneva: 2011.
-
- Snow RW, Korenromp EL, Gouws E. Pediatric mortality in Africa: plasmodium falciparum malaria as a cause or risk? Am. J. Trop. Med. Hyg. 2004;71:16–24. - PubMed
-
- Berkley JA, Bejon P, Mwangi T, Gwer S, Maitland K, Williams TN, Mohammed S, Osier F, Kinyanjui S, Fegan G, Lowe BS, English M, Peshu N, Marsh K, Newton CR. HIV infection, malnutrition, and invasive bacterial infection among children with severe malaria. Clin. Infect. Dis. 2009;49:336–343. - PMC - PubMed
-
- Scott JA, Berkley JA, Mwangi I, Ochola L, Uyoga S, Macharia A, Ndila C, Lowe BS, Mwarumba S, Bauni E, Marsh K, Williams TN. Relation between falciparum malaria and bacteraemia in Kenyan children: a population-based, case-control study and a longitudinal study. Lancet. 2011;378:1316–1323. - PMC - PubMed
-
- Bronzan RN, Taylor TE, Mwenechanya J, Tembo M, Kayira K, Bwanaisa L, Njobvu A, Kondowe W, Chalira C, Walsh AL, Phiri A, Wilson LK, Molyneux ME, Graham SM. Bacteremia in Malawian children with severe malaria: prevalence, etiology, HIV coinfection, and outcome. J. Infect. Dis. 2007;195:895–904. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials