Ixodes scapularis saliva mitigates inflammatory cytokine secretion during Anaplasma phagocytophilum stimulation of immune cells
- PMID: 23050849
- PMCID: PMC3503595
- DOI: 10.1186/1756-3305年5月22日9
Ixodes scapularis saliva mitigates inflammatory cytokine secretion during Anaplasma phagocytophilum stimulation of immune cells
Abstract
Background: Ixodes scapularis saliva enables the transmission of infectious agents to the mammalian host due to its immunomodulatory, anesthetic and anti-coagulant properties. However, how I. scapularis saliva influences host cytokine secretion in the presence of the obligate intracellular rickettsial pathogen Anaplasma phagocytophilum remains elusive.
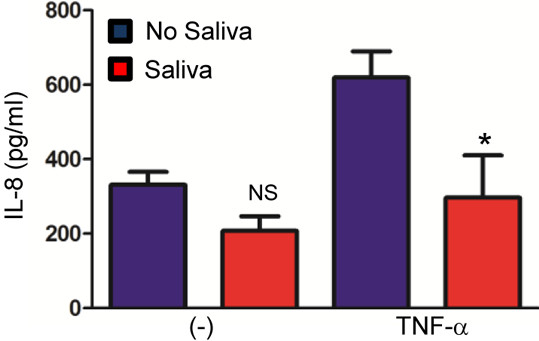
Methods: Bone marrow derived macrophages (BMDMs) were stimulated with pathogen associated molecular patterns (PAMPs) and A. phagocytophilum. Cytokine secretion was measured in the presence and absence of I. scapularis saliva. Human peripheral blood mononuclear cells (PBMCs) were also stimulated with Tumor Necrosis Factor (TNF)-α in the presence and absence of I. scapularis saliva and interleukin (IL)-8 was measured.
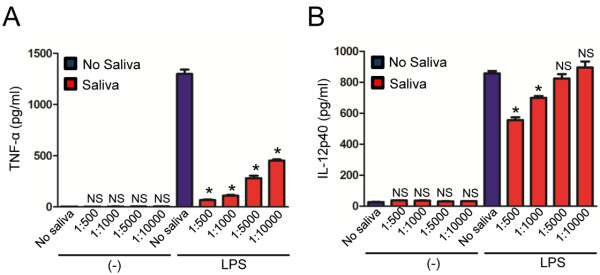
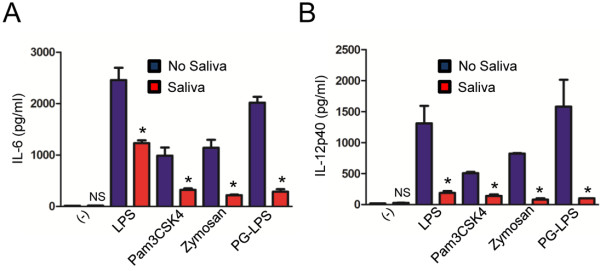
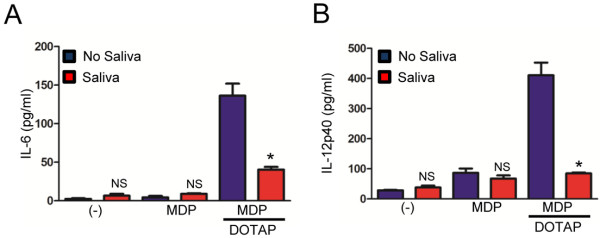
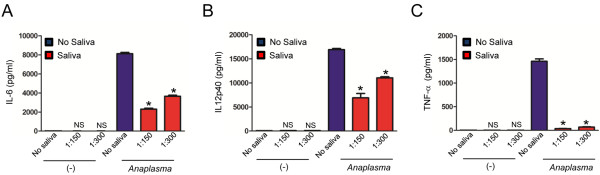
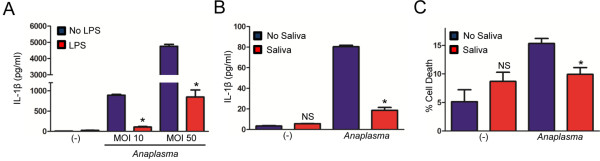
Results: I. scapularis saliva inhibits inflammatory cytokine secretion by macrophages during stimulation of Toll-like (TLR) and Nod-like receptor (NLR) signaling pathways. The effect of I. scapularis saliva on immune cells is not restricted to murine macrophages because decreasing levels of interleukin (IL)-8 were observed after TNF-α stimulation of human peripheral blood mononuclear cells. I. scapularis saliva also mitigates pro-inflammatory cytokine response by murine macrophages during challenge with A. phagocytophilum.
Conclusions: These findings suggest that I. scapularis may inhibit inflammatory cytokine secretion during rickettsial transmission at the vector-host interface.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Valenzuela JG. Exploring tick saliva: from biochemistry to ‘sialomes’ and functional genomics. Parasitology. 2004;129(Suppl):S83–S94. - PubMed