The life cycle of Trypanosoma (Nannomonas) congolense in the tsetse fly
- PMID: 22676292
- PMCID: PMC3384477
- DOI: 10.1186/1756-3305年5月10日9
The life cycle of Trypanosoma (Nannomonas) congolense in the tsetse fly
Abstract
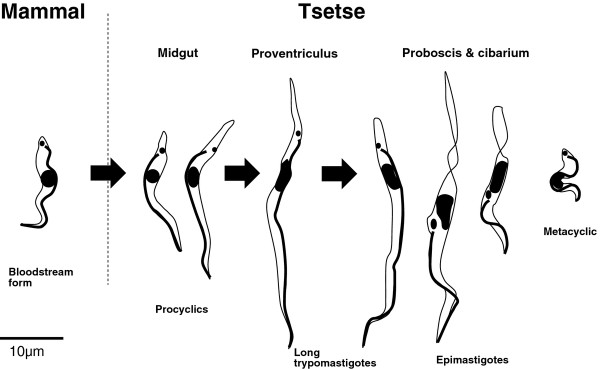
Background: The tsetse-transmitted African trypanosomes cause diseases of importance to the health of both humans and livestock. The life cycles of these trypanosomes in the fly were described in the last century, but comparatively few details are available for Trypanosoma (Nannomonas) congolense, despite the fact that it is probably the most prevalent and widespread pathogenic species for livestock in tropical Africa. When the fly takes up bloodstream form trypanosomes, the initial establishment of midgut infection and invasion of the proventriculus is much the same in T. congolense and T. brucei. However, the developmental pathways subsequently diverge, with production of infective metacyclics in the proboscis for T. congolense and in the salivary glands for T. brucei. Whereas events during migration from the proventriculus are understood for T. brucei, knowledge of the corresponding developmental pathway in T. congolense is rudimentary. The recent publication of the genome sequence makes it timely to re-investigate the life cycle of T. congolense.
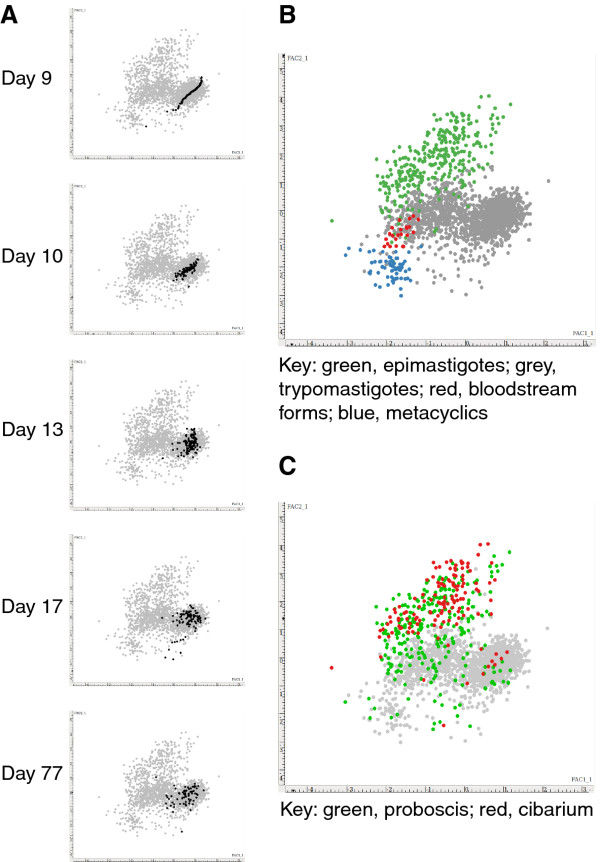
Methods: Experimental tsetse flies were fed an initial bloodmeal containing T. congolense strain 1/148 and dissected 2 to 78 days later. Trypanosomes recovered from the midgut, proventriculus, proboscis and cibarium were fixed and stained for digital image analysis. Trypanosomes contained in spit samples from individually caged flies were analysed similarly. Mensural data from individual trypanosomes were subjected to principal components analysis.
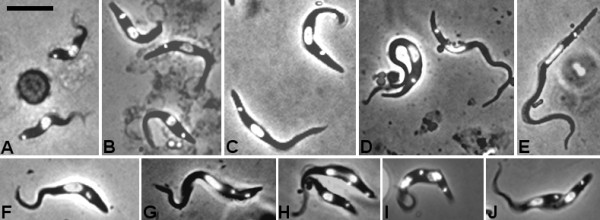
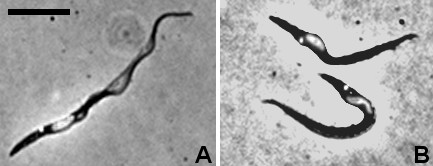
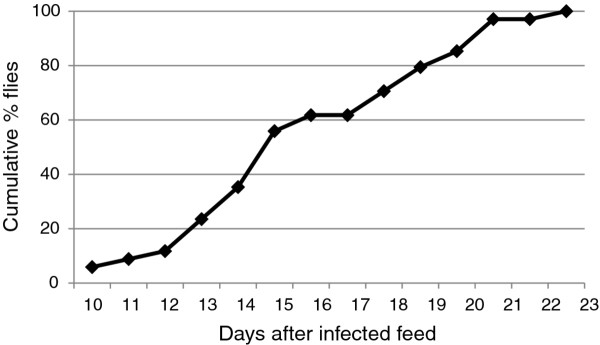
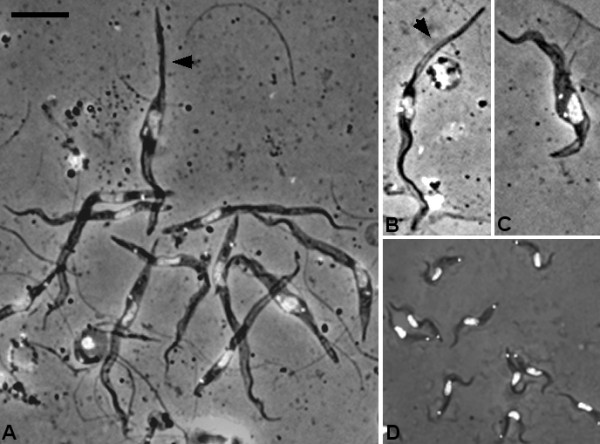
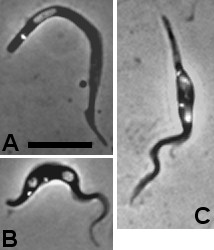
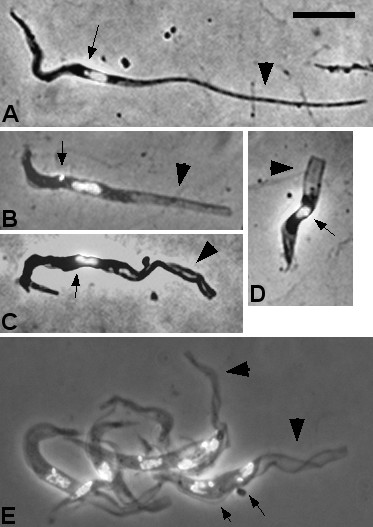
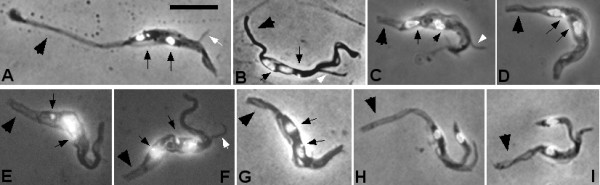
Results: Flies were more susceptible to infection with T. congolense than T. brucei; a high proportion of flies infected with T. congolense established a midgut and subsequent proboscis infection, whereas many T. brucei infections were lost in the migration from foregut to salivary glands. In T. congolense, trypomastigotes ceased division in the proventriculus and became uniform in size. The trypanosomes retained trypomastigote morphology during migration via the foregut to the mouthparts and we confirmed that the trypomastigote-epimastigote transition occurred in the proboscis. We found no equivalent to the asymmetric division stage in T. brucei that mediates transition of proventricular trypomastigotes to epimastigotes. In T. congolense extremely long epimastigotes with remarkably elongated posterior ends were observed in both the proboscis and cibarium; no difference was found in the developmental stages in these two organs. Dividing trypomastigotes and epimastigotes were recovered from the proboscis, some of which were in transition from trypomastigote to epimastigote and vice versa. It remains uncertain whether these morphological transitions are mediated by cell division, since we also found non-dividing cells with a variously positioned, juxta-nuclear kinetoplast.
Conclusions: We have presented a detailed description of the life cycle of T. congolense in its tsetse fly vector. During development in the fly T. congolense shares a common migratory pathway with its close relative T. brucei, culminating in the production of small metacyclic trypanosomes that can be inoculated with the saliva. Despite this outward similarity in life cycle, the transitional developmental stages in the foregut and mouthparts are remarkably different in the two trypanosome species.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Stephen LE. Trypanosomiasis, a veterinary perspective. Pergamon Press, Oxford; 1986.
-
- Hoare CA. The Trypanosomes of Mammals. Blackwell Scientific Publications, Oxford; 1972.
-
- Matthews KR, Gull K. Commitment to differentiation and cell cycle re-entry are coincident but separable events in the transformation of African trypanosomes from their bloodstream to their insect form. J Cell Sci. 1997;110:2609–2618. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources