Inducible nitric oxide synthase in heart tissue and nitric oxide in serum of Trypanosoma cruzi-infected rhesus monkeys: association with heart injury
- PMID: 22590660
- PMCID: PMC3348164
- DOI: 10.1371/journal.pntd.0001644
Inducible nitric oxide synthase in heart tissue and nitric oxide in serum of Trypanosoma cruzi-infected rhesus monkeys: association with heart injury
Abstract
Background: The factors contributing to chronic Chagas' heart disease remain unknown. High nitric oxide (NO) levels have been shown to be associated with cardiomyopathy severity in patients. Further, NO produced via inducible nitric oxide synthase (iNOS/NOS2) is proposed to play a role in Trypanosoma cruzi control. However, the participation of iNOS/NOS2 and NO in T. cruzi control and heart injury has been questioned. Here, using chronically infected rhesus monkeys and iNOS/NOS2-deficient (Nos2(-/-)) mice we explored the participation of iNOS/NOS2-derived NO in heart injury in T. cruzi infection.
Methodology: Rhesus monkeys and C57BL/6 and Nos2(-/-) mice were infected with the Colombian T. cruzi strain. Parasite DNA was detected by polymerase chain reaction, T. cruzi antigens and iNOS/NOS2(+) cells were immunohistochemically detected in heart sections and NO levels in serum were determined by Griess reagent. Heart injury was assessed by electrocardiogram (ECG), echocardiogram (ECHO), creatine kinase heart isoenzyme (CK-MB) activity levels in serum and connexin 43 (Cx43) expression in the cardiac tissue.
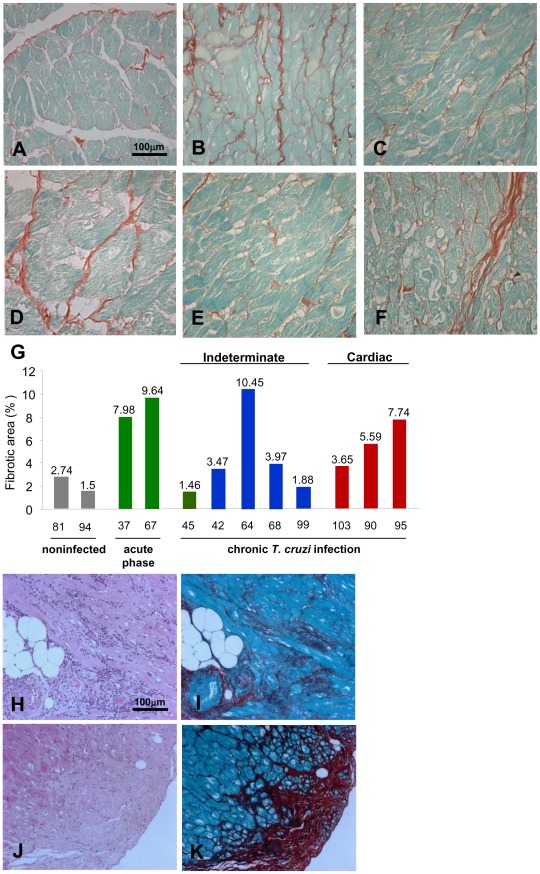
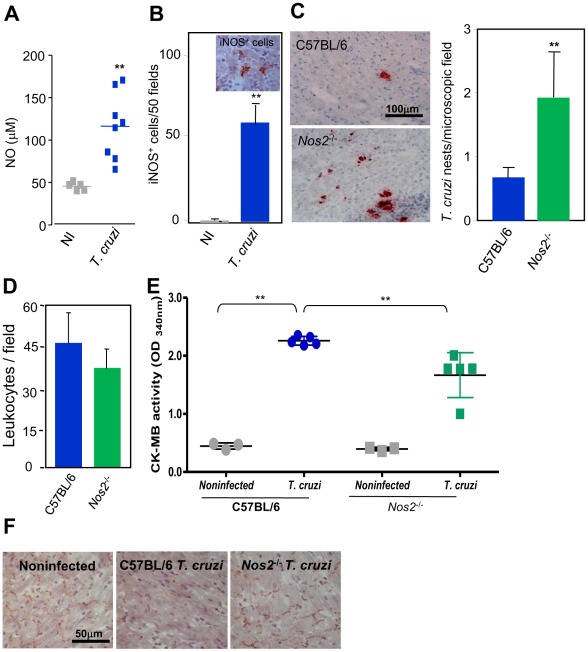
Results: Chronically infected monkeys presented conduction abnormalities, cardiac inflammation and fibrosis, which resembled the spectrum of human chronic chagasic cardiomyopathy (CCC). Importantly, chronic myocarditis was associated with parasite persistence. Moreover, Cx43 loss and increased CK-MB activity levels were primarily correlated with iNOS/NOS2(+) cells infiltrating the cardiac tissue and NO levels in serum. Studies in Nos2(-/-) mice reinforced that the iNOS/NOS2-NO pathway plays a pivotal role in T. cruzi-elicited cardiomyocyte injury and in conduction abnormalities that were associated with Cx43 loss in the cardiac tissue.
Conclusion: T. cruzi-infected rhesus monkeys reproduce features of CCC. Moreover, our data support that in T. cruzi infection persistent parasite-triggered iNOS/NOS2 in the cardiac tissue and NO overproduction might contribute to CCC severity, mainly disturbing of the molecular pathway involved in electrical synchrony. These findings open a new avenue for therapeutic tools in Chagas' heart disease.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Rassi A, Jr, Rassi A, Marin-Neto JA. Chagas disease. Lancet. 2010;375:1388–1402. - PubMed
-
- Kierszenbaum F. Where do we stand on the autoimmunity hypothesis of Chagas disease? Trends Parasitol. 2005;21:513–516. - PubMed
-
- Higuchi ML, Gutierrez PS, Aiello VD, Palomino S, Bocchi E, et al. Immunohistochemical characterization of infiltrating cells in human chronic chagasic myocarditis: comparison with myocardial rejection process. Virchows Arch A Pathol Anat Histopathol. 1993;423:157–160. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous