Crimean-Congo hemorrhagic fever virus nucleoprotein reveals endonuclease activity in bunyaviruses
- PMID: 22421137
- PMCID: PMC3324003
- DOI: 10.1073/pnas.1200808109
Crimean-Congo hemorrhagic fever virus nucleoprotein reveals endonuclease activity in bunyaviruses
Abstract
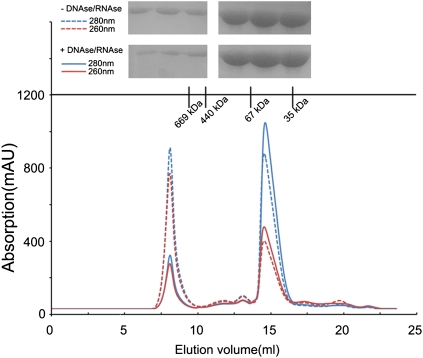
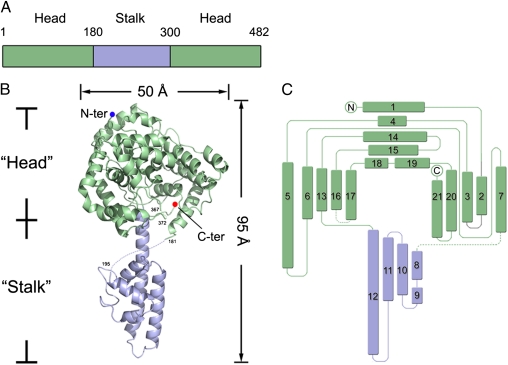
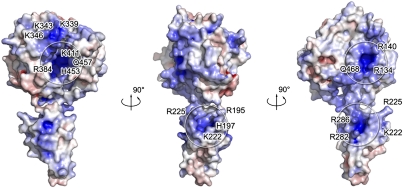
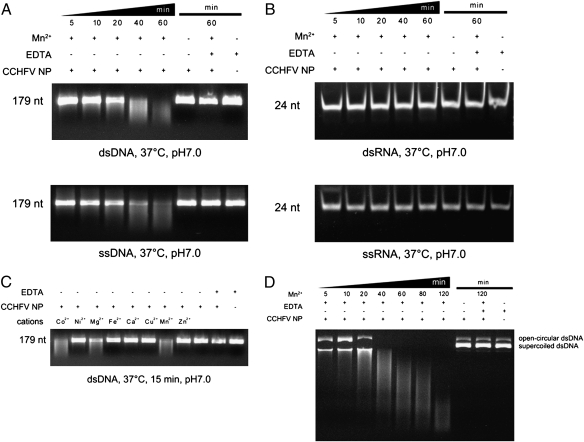
Crimean-Congo hemorrhagic fever virus (CCHFV), a virus with high mortality in humans, is a member of the genus Nairovirus in the family Bunyaviridae, and is a causative agent of severe hemorrhagic fever (HF). It is classified as a biosafety level 4 pathogen and a potential bioterrorism agent due to its aerosol infectivity and its ability to cause HF outbreaks with high case fatality (∼30%). However, little is known about the structural features and function of nucleoproteins (NPs) in the Bunyaviridae, especially in CCHFV. Here we report a 2.3-Å resolution crystal structure of the CCHFV nucleoprotein. The protein has a racket-shaped overall structure with distinct "head" and "stalk" domains and differs significantly with NPs reported so far from other negative-sense single-stranded RNA viruses. Furthermore, CCHFV NP shows a distinct metal-dependent DNA-specific endonuclease activity. Single residue mutations in the predicted active site resulted in a significant reduction in the observed endonuclease activity. Our results present a new folding mechanism and function for a negative-strand RNA virus nucleoprotein, extend our structural insight into bunyavirus NPs, and provide a potential target for antiviral drug development to treat CCHFV infection.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Flick R, Whitehouse CA. Crimean-Congo hemorrhagic fever virus. Curr Mol Med. 2005;5:753–760. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous