Valosin-containing protein (VCP/p97) is required for poliovirus replication and is involved in cellular protein secretion pathway in poliovirus infection
- PMID: 22379090
- PMCID: PMC3347272
- DOI: 10.1128/JVI.00114-12
Valosin-containing protein (VCP/p97) is required for poliovirus replication and is involved in cellular protein secretion pathway in poliovirus infection
Abstract
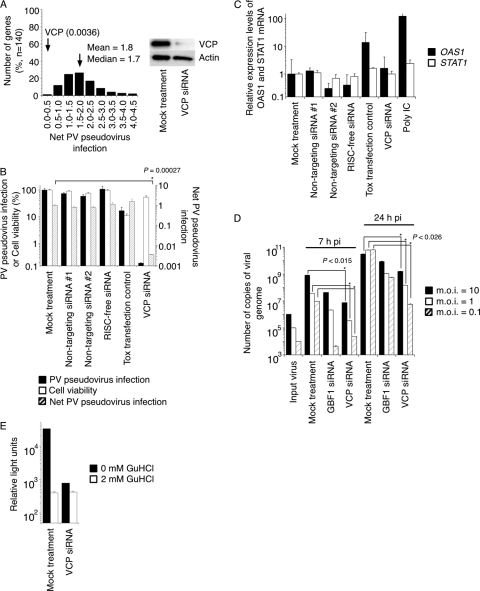
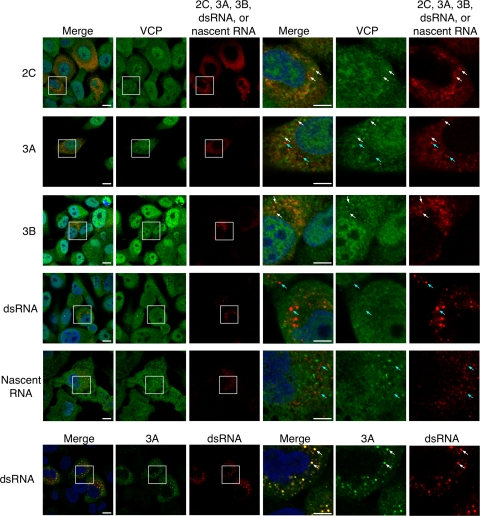
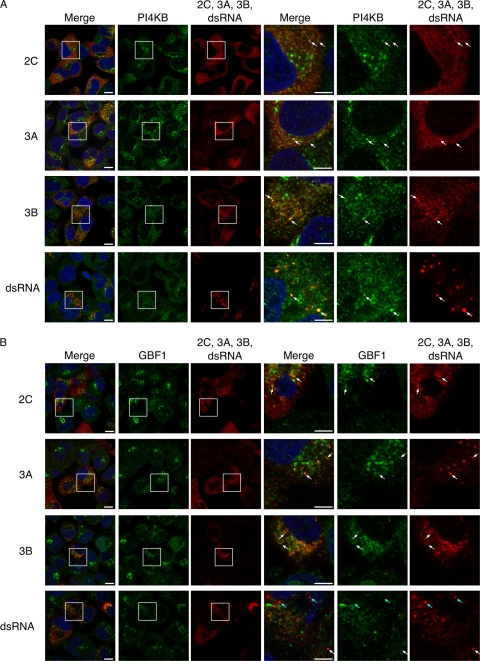
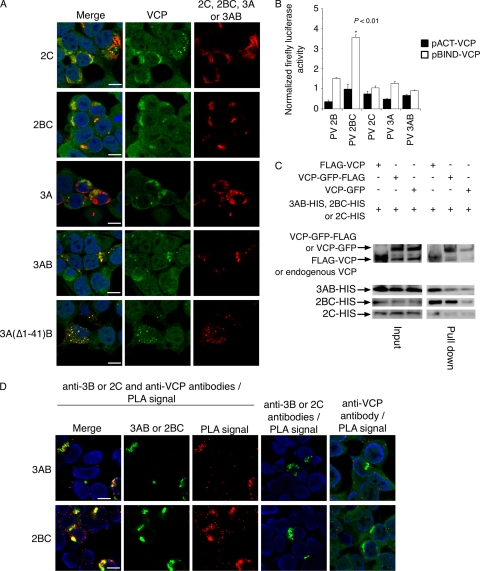
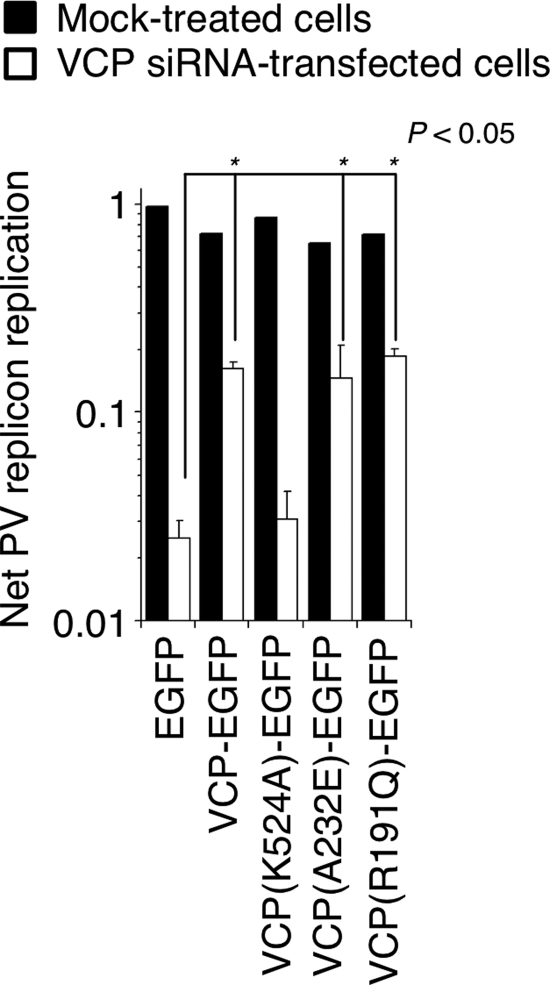
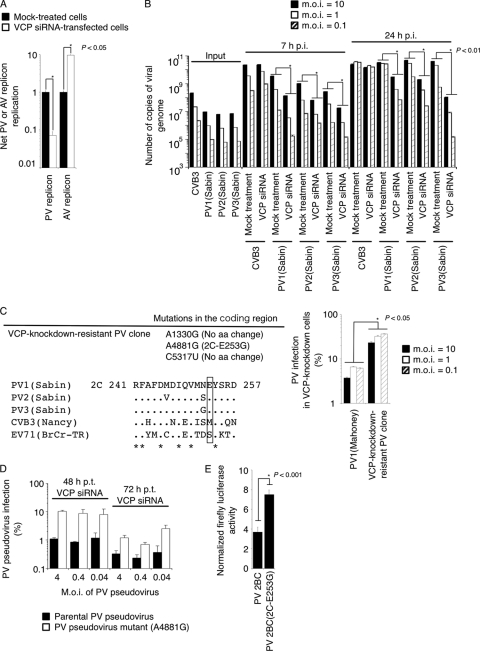
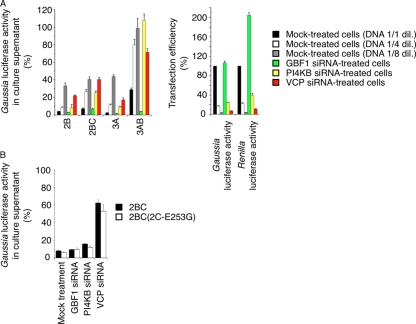
Poliovirus (PV) modifies membrane-trafficking machinery in host cells for its viral RNA replication. To date, ARF1, ACBD3, BIG1/BIG2, GBF1, RTN3, and PI4KB have been identified as host factors of enterovirus (EV), including PV, involved in membrane traffic. In this study, we performed small interfering RNA (siRNA) screening targeting membrane-trafficking genes for host factors required for PV replication. We identified valosin-containing protein (VCP/p97) as a host factor of PV replication required after viral protein synthesis, and its ATPase activity was essential for PV replication. VCP colocalized with viral proteins 2BC/2C and 3AB/3B in PV-infected cells and showed an interaction with 2BC and 3AB but not with 2C and 3A. Knockdown of VCP did not suppress the replication of coxsackievirus B3 or Aichi virus. A VCP-knockdown-resistant PV mutant had an A4881G (a mutation of E253G in 2C) mutation, which is known as a determinant of a secretion inhibition-negative phenotype. However, knockdown of VCP did not affect the inhibition of cellular protein secretion caused by overexpression of each individual viral protein. These results suggested that VCP is a host factor required for viral RNA replication of PV among membrane-trafficking proteins and provides a novel link between cellular protein secretion and viral RNA replication.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Arita M, Nagata N, Sata T, Miyamura T, Shimizu H. 2006. Quantitative analysis of poliomyelitis-like paralysis in mice induced by a poliovirus replicon. J. Gen. Virol. 87:3317–3327 - PubMed
-
- Arita M, Takebe Y, Wakita T, Shimizu H. 2010. A bifunctional anti-enterovirus compound that inhibits replication and early stage of enterovirus 71 infection. J. Gen. Virol. 91:2734–2744 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous