A periodically-forced mathematical model for the seasonal dynamics of malaria in mosquitoes
- PMID: 22218880
- PMCID: PMC3339865
- DOI: 10.1007/s11538-011-9710-0
A periodically-forced mathematical model for the seasonal dynamics of malaria in mosquitoes
Abstract
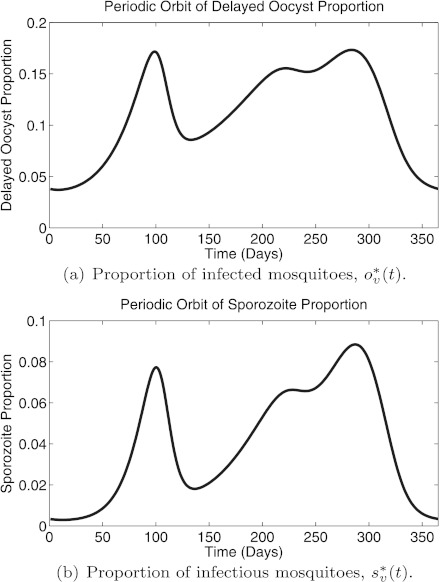
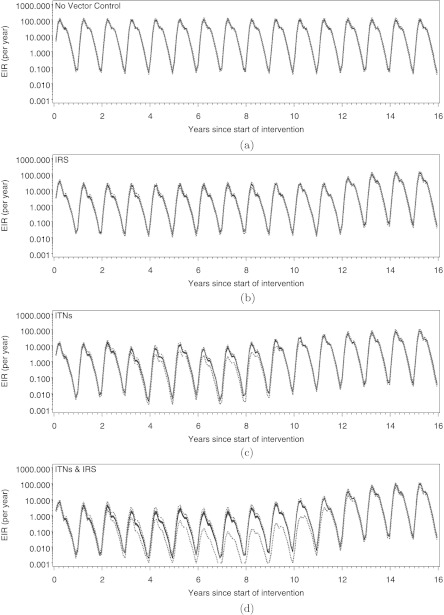
We describe and analyze a periodically-forced difference equation model for malaria in mosquitoes that captures the effects of seasonality and allows the mosquitoes to feed on a heterogeneous population of hosts. We numerically show the existence of a unique globally asymptotically stable periodic orbit and calculate periodic orbits of field-measurable quantities that measure malaria transmission. We integrate this model with an individual-based stochastic simulation model for malaria in humans to compare the effects of insecticide-treated nets (ITNs) and indoor residual spraying (IRS) in reducing malaria transmission, prevalence, and incidence. We show that ITNs are more effective than IRS in reducing transmission and prevalence though IRS would achieve its maximal effects within 2 years while ITNs would need two mass distribution campaigns over several years to do so. Furthermore, the combination of both interventions is more effective than either intervention alone. However, although these interventions reduce transmission and prevalence, they can lead to increased clinical malaria; and all three malaria indicators return to preintervention levels within 3 years after the interventions are withdrawn.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Anderson R. M., May R. M. Infectious diseases of humans: dynamics and control. Oxford: Oxford Unversity Press; 1991.
-
- Aron J. L. Mathematical modeling of immunity to malaria. Math. Biosci. 1988;90:385–396. doi: 10.1016/0025-5564(88)90076-4. - DOI
-
- Aron J. L., May R. M. The population dynamics of malaria. In: Anderson R. M., editor. The population dynamics of infectious disease: theory and applications. London: Chapman and Hall; 1982. pp. 139–179.
-
- Charlwood J. D., Smith T., Billingsley P. F., Takken W., Lyimo E. O. K., Meuwissen J. H. E. T. Survival and infection probabilities of anthropophagic anophelines from an area of high prevalence of Plasmodium falciparum in humans. Bull. Entomol. Res. 1997;87:445–453. doi: 10.1017/S0007485300041304. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical