Genomic maps of long noncoding RNA occupancy reveal principles of RNA-chromatin interactions
- PMID: 21963238
- PMCID: PMC3249421
- DOI: 10.1016/j.molcel.2011年08月02日7
Genomic maps of long noncoding RNA occupancy reveal principles of RNA-chromatin interactions
Abstract
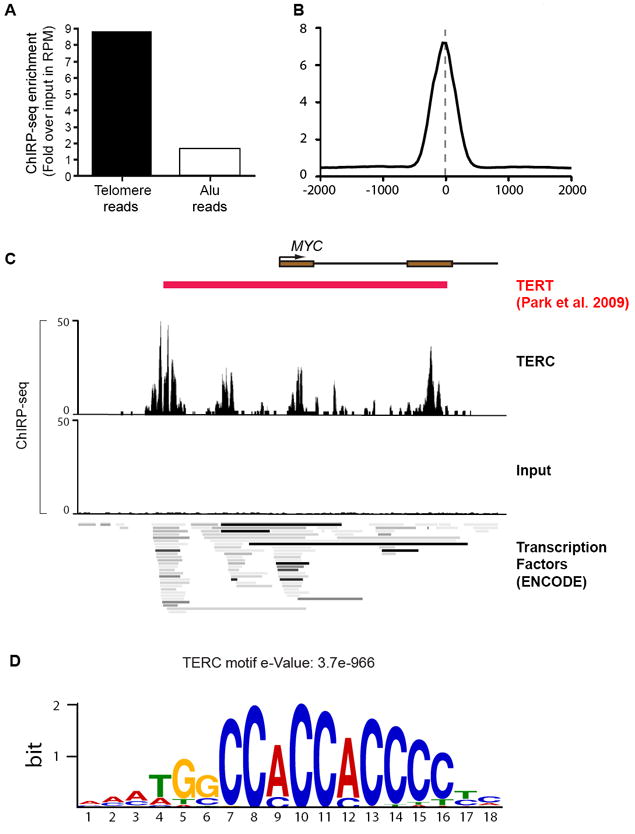
Long noncoding RNAs (lncRNAs) are key regulators of chromatin state, yet the nature and sites of RNA-chromatin interaction are mostly unknown. Here we introduce Chromatin Isolation by RNA Purification (ChIRP), where tiling oligonucleotides retrieve specific lncRNAs with bound protein and DNA sequences, which are enumerated by deep sequencing. ChIRP-seq of three lncRNAs reveal that RNA occupancy sites in the genome are focal, sequence-specific, and numerous. Drosophila roX2 RNA occupies male X-linked gene bodies with increasing tendency toward the 3' end, peaking at CES sites. Human telomerase RNA TERC occupies telomeres and Wnt pathway genes. HOTAIR lncRNA preferentially occupies a GA-rich DNA motif to nucleate broad domains of Polycomb occupancy and histone H3 lysine 27 trimethylation. HOTAIR occupancy occurs independently of EZH2, suggesting the order of RNA guidance of Polycomb occupancy. ChIRP-seq is generally applicable to illuminate the intersection of RNA and chromatin with newfound precision genome wide.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Comment in
-
RNA: Genome-wide views of long non-coding RNAs.Muers M. Muers M. Nat Rev Genet. 2011 Oct 12;12(11):742. doi: 10.1038/nrg3088. Nat Rev Genet. 2011. PMID: 21989130 No abstract available.
-
Noncoding RNA's genomic hangouts.Nawy T. Nawy T. Nat Methods. 2011 Dec;8(12):1000. doi: 10.1038/nmeth.1787. Nat Methods. 2011. PMID: 22238778 No abstract available.
References
-
- Bailey TL, Elkan C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc Int Conf Intell Syst Mol Biol. 1994;2:28–36. - PubMed
-
- Carter D, Chakalova L, Osborne CS, Dai YF, Fraser P. Long-range chromatin regulatory interactions in vivo. Nature genetics. 2002;32:623–626. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases