Avoiding DEET through insect gustatory receptors
- PMID: 20797533
- PMCID: PMC2929391
- DOI: 10.1016/j.neuron.201007006
Avoiding DEET through insect gustatory receptors
Abstract
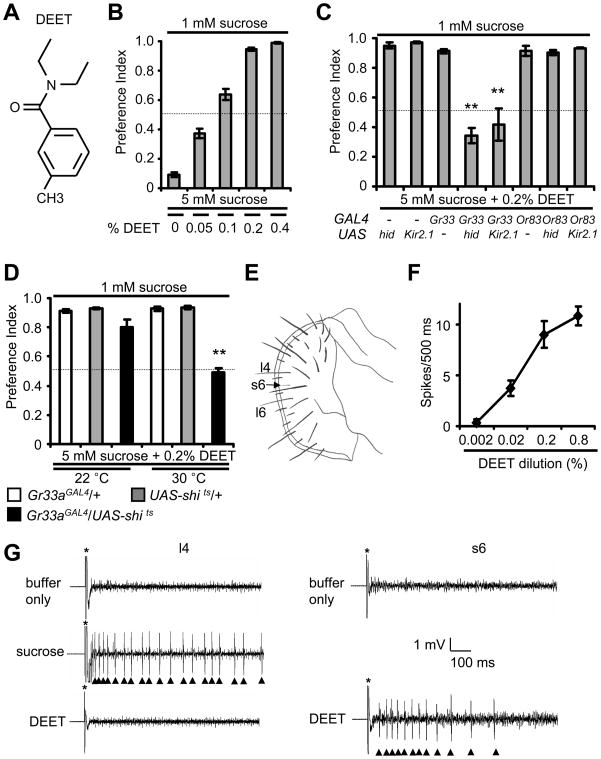
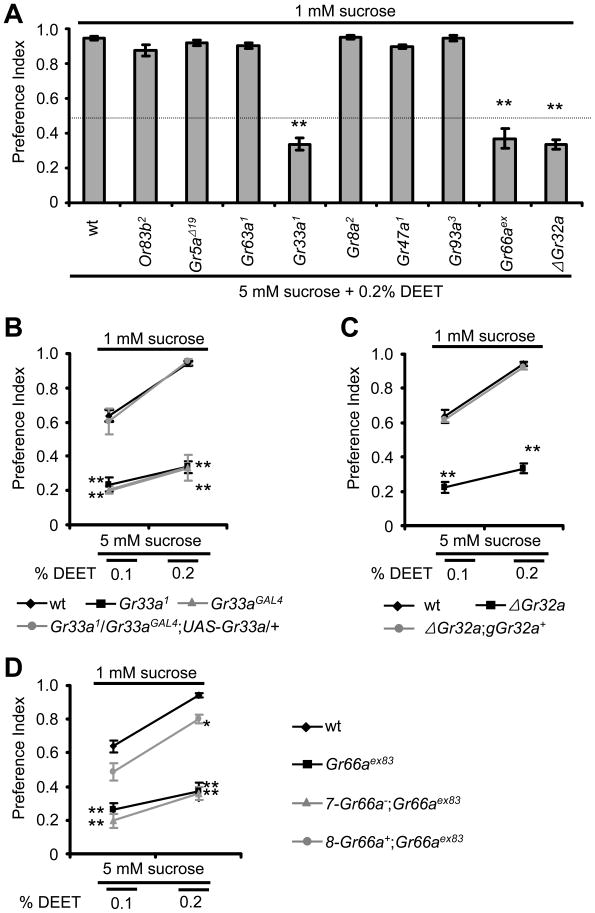
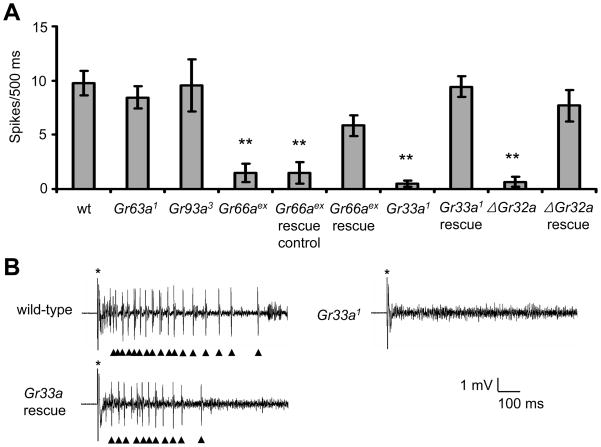
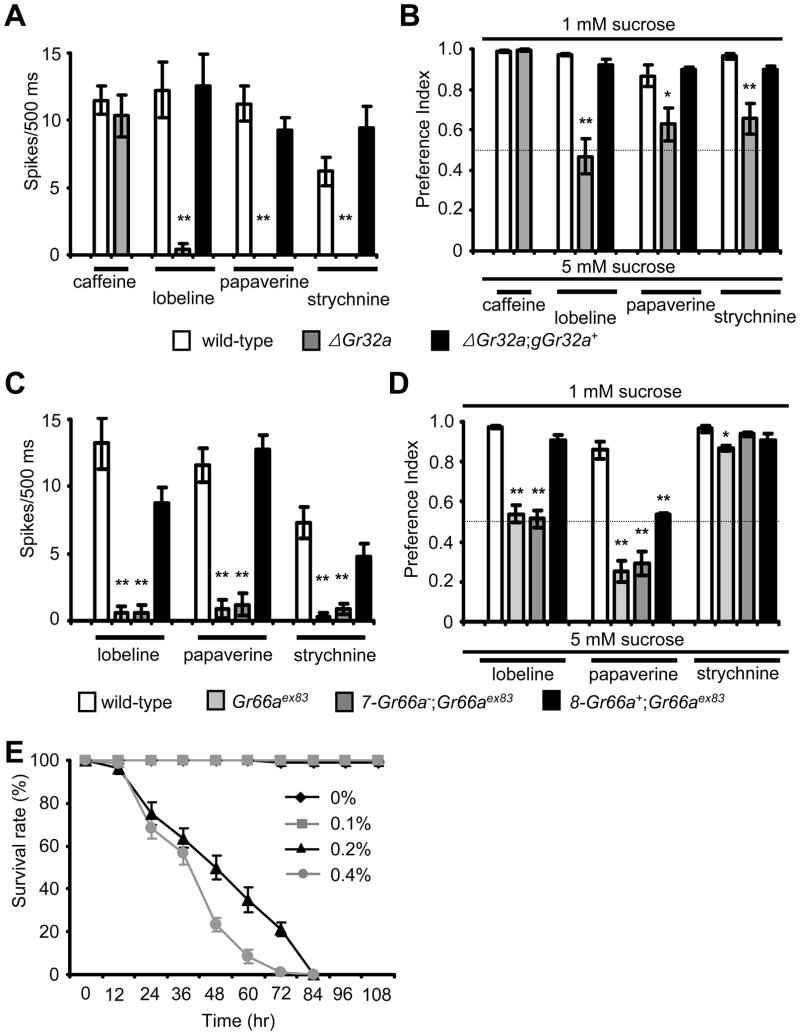
DEET is the most widely used insect repellent worldwide. In Drosophila olfactory receptor neurons (ORNs), DEET is detected through a mechanism employing the olfactory receptor, OR83b. However, it is controversial as to whether ORNs respond directly to DEET or whether DEET blocks the response to attractive odors. Here, we showed that DEET suppressed feeding behavior in Drosophila, and this effect was mediated by gustatory receptor neurons (GRNs). DEET was potent in suppressing feeding as <0.1% DEET elicited aversive behavior. Inhibition of feeding required multiple gustatory receptors (GRs) expressed in inhibitory GRNs. DEET stimulated action potentials in GRNs that respond to aversive compounds, and this response was lost in the Gr32a, Gr33a, and Gr66a mutants. Since 0.02% DEET elicited action potentials, we conclude that DEET directly activates of GRNs. We suggest that the effectiveness of DEET in pest control owes to its dual action in inducing avoidance simultaneously via GRNs and ORNs.
2010 Elsevier Inc. All rights reserved.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Ditzen M, Pellegrino M, Vosshall LB. Insect odorant receptors are molecular targets of the insect repellent DEET. Science. 2008;319:1838–1842. - PubMed
-
- Dogan EB, Ayres JW, Rossignol PA. Behavioural mode of action of deet: inhibition of lactic acid attraction. Med Vet Entomol. 1999;13:97–100. - PubMed
-
- Hiroi M, Marion-Poll F, Tanimura T. Differentiated response to sugars among labellar chemosensilla in Drosophila. Zoolog Sci. 2002;19:1009–1018. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases