Ablation of succinate production from glucose metabolism in the procyclic trypanosomes induces metabolic switches to the glycerol 3-phosphate/dihydroxyacetone phosphate shuttle and to proline metabolism
- PMID: 20702405
- PMCID: PMC2952232
- DOI: 10.1074/jbc.M110.124917
Ablation of succinate production from glucose metabolism in the procyclic trypanosomes induces metabolic switches to the glycerol 3-phosphate/dihydroxyacetone phosphate shuttle and to proline metabolism
Abstract
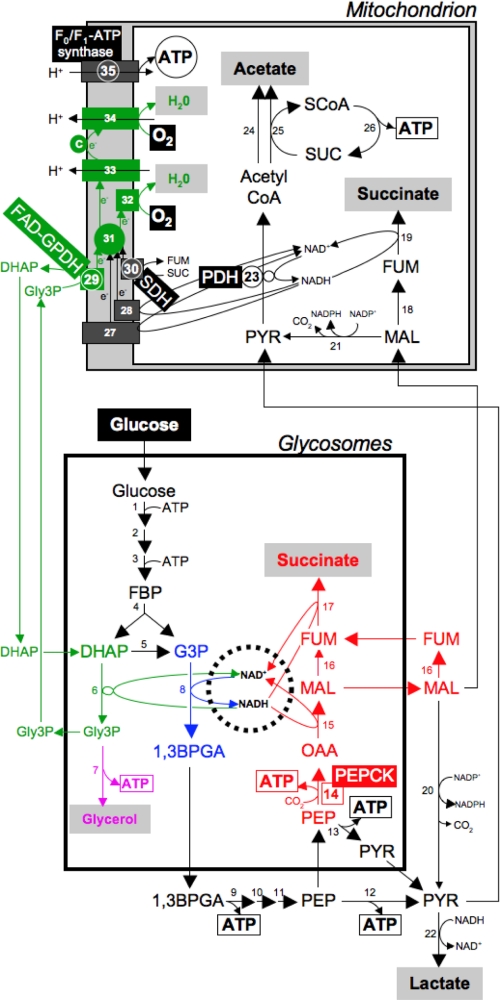
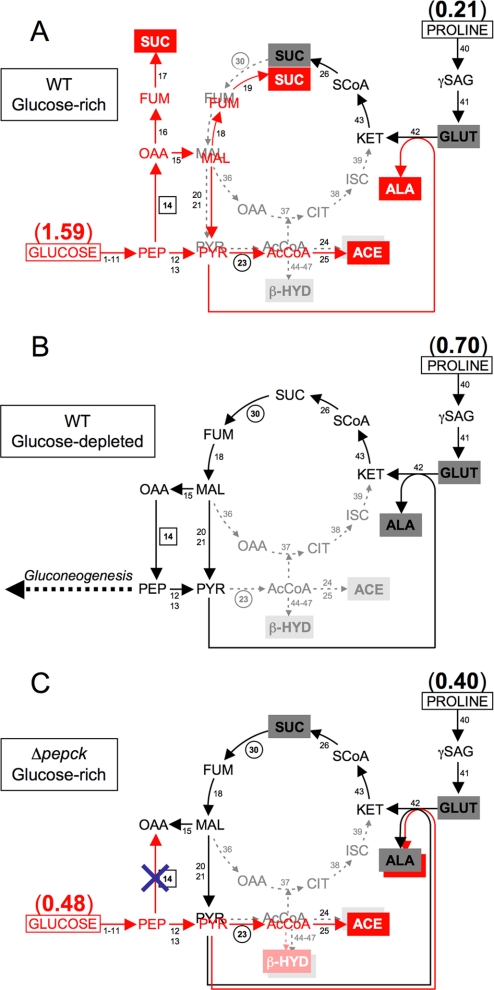
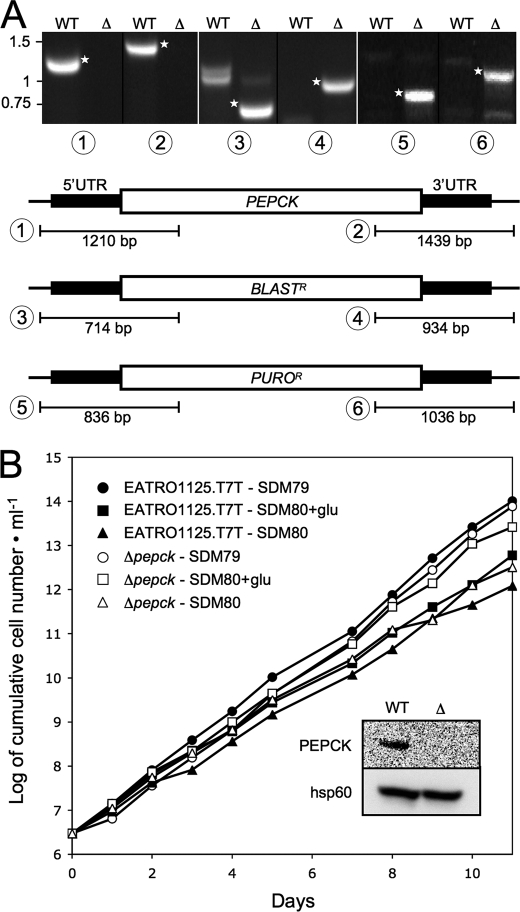
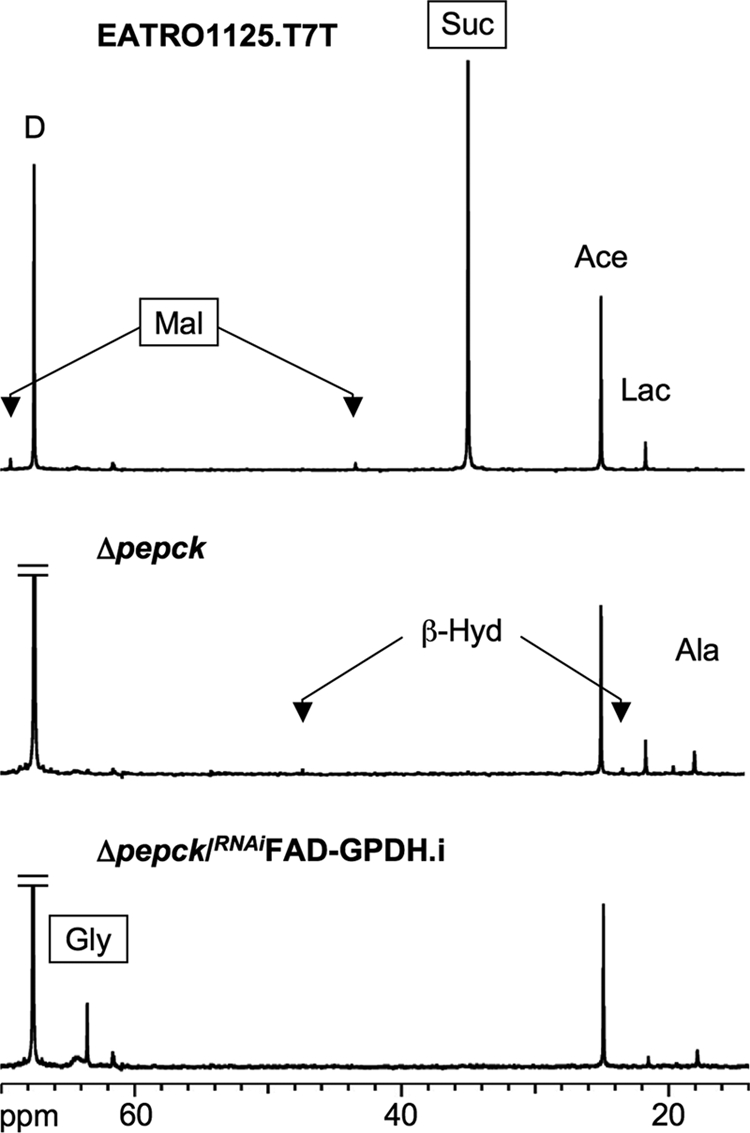
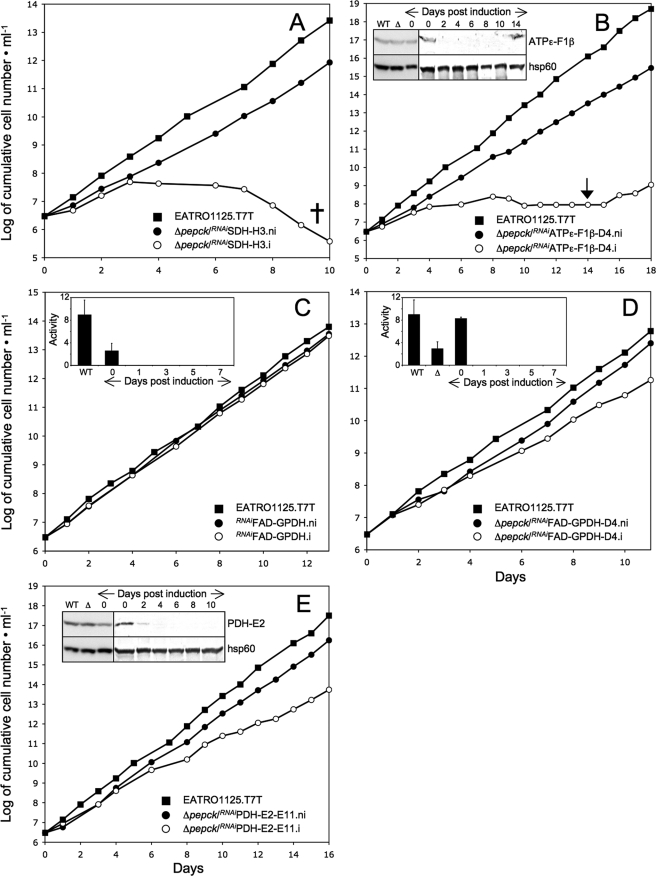
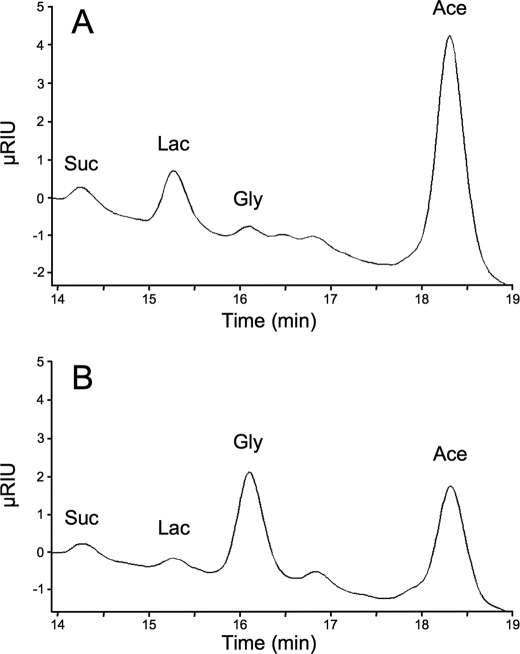
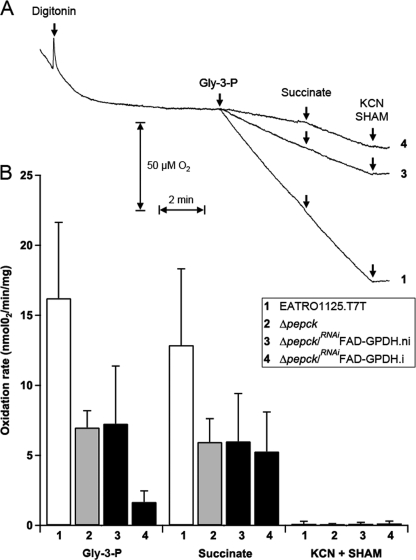
Trypanosoma brucei is a parasitic protist that undergoes a complex life cycle during transmission from its mammalian host (bloodstream forms) to the midgut of its insect vector (procyclic form). In both parasitic forms, most glycolytic steps take place within specialized peroxisomes, called glycosomes. Here, we studied metabolic adaptations in procyclic trypanosome mutants affected in their maintenance of the glycosomal redox balance. T. brucei can theoretically use three strategies to maintain the glycosomal NAD(+)/NADH balance as follows: (i) the glycosomal succinic fermentation branch; (ii) the glycerol 3-phosphate (Gly-3-P)/dihydroxyacetone phosphate (DHAP) shuttle that transfers reducing equivalents to the mitochondrion; and (iii) the glycosomal glycerol production pathway. We showed a hierarchy in the use of these glycosomal NADH-consuming pathways by determining metabolic perturbations and adaptations in single and double mutant cell lines using a combination of NMR, ion chromatography-MS/MS, and HPLC approaches. Although functional, the Gly-3-P/DHAP shuttle is primarily used when the preferred succinate fermentation pathway is abolished in the Δpepck knock-out mutant cell line. In the absence of these two pathways (Δpepck/(RNAi)FAD-GPDH.i mutant), glycerol production is used but with a 16-fold reduced glycolytic flux. In addition, the Δpepck mutant cell line shows a 3.3-fold reduced glycolytic flux compensated by an increase of proline metabolism. The inability of the Δpepck mutant to maintain a high glycolytic flux demonstrates that the Gly-3-P/DHAP shuttle is not adapted to the procyclic trypanosome context. In contrast, this shuttle was shown earlier to be the only way used by the bloodstream forms of T. brucei to sustain their high glycolytic flux.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Barrett M. P., Burchmore R. J., Stich A., Lazzari J. O., Frasch A. C., Cazzulo J. J., Krishna S. (2003) Lancet 362, 1469–1480 - PubMed
-
- Michels P. A., Bringaud F., Herman M., Hannaert V. (2006) Biochim. Biophys. Acta 1763, 1463–1477 - PubMed
-
- Cross G. A., Klein R. A., Linstead D. J. (1975) Parasitology 71, 311–326 - PubMed
-
- Coustou V., Besteiro S., Biran M., Diolez P., Bouchaud V., Voisin P., Michels P. A., Canioni P., Baltz T., Bringaud F. (2003) J. Biol. Chem. 278, 49625–49635 - PubMed
-
- Fairlamb A. H., Opperdoes F. R. (1986) in Carbohydrate Metabolism in Cultured Cells (Morgan M. J. ed) pp. 183–224, Plenum Publishing Corp., New York
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources