Cryptococcal cell morphology affects host cell interactions and pathogenicity
- PMID: 20585559
- PMCID: PMC2887476
- DOI: 10.1371/journal.ppat.1000953
Cryptococcal cell morphology affects host cell interactions and pathogenicity
Erratum in
- PLoS Pathog. 2010;6(6). doi: 10.1371/annotation/1b59fd9e-9ac9-4ea8-a083-14c413c80b03
Abstract
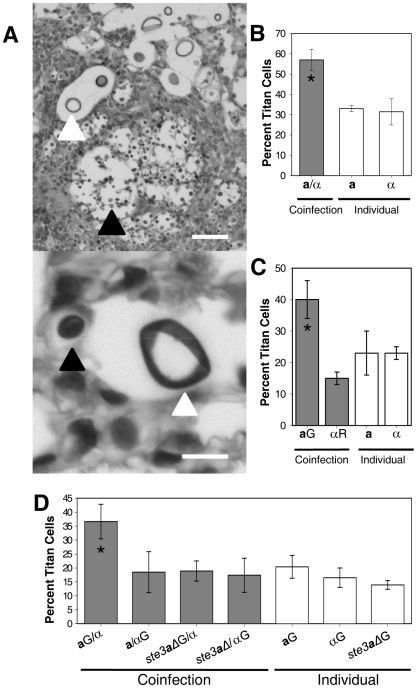
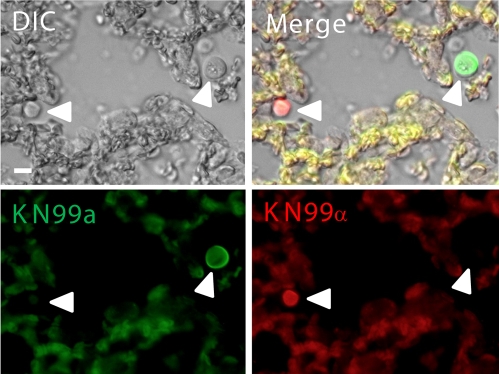
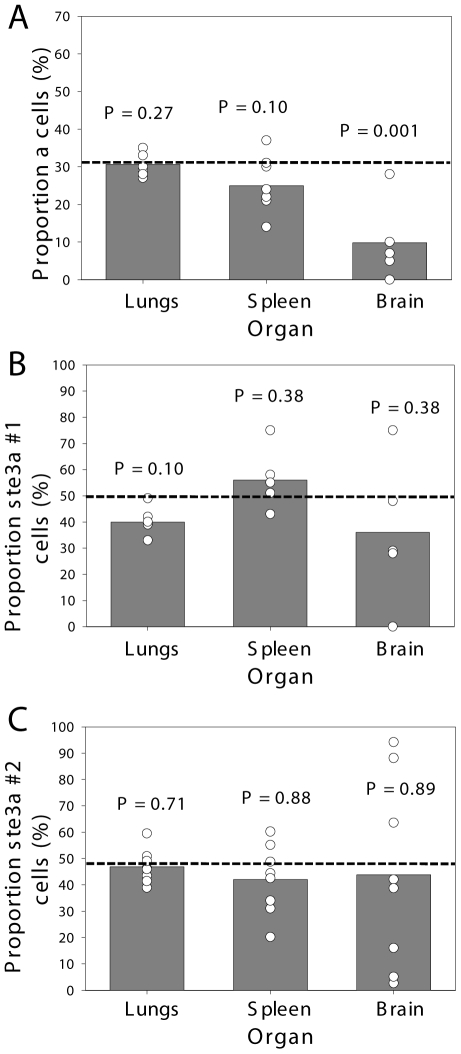
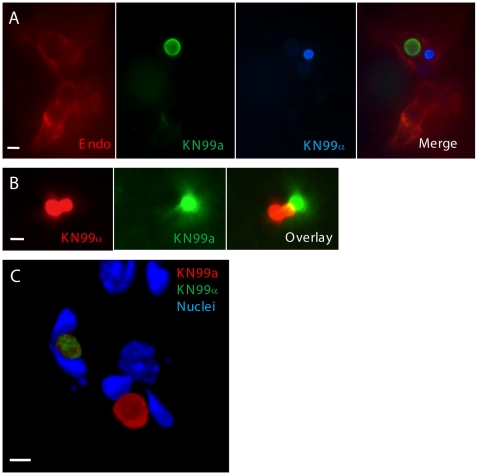
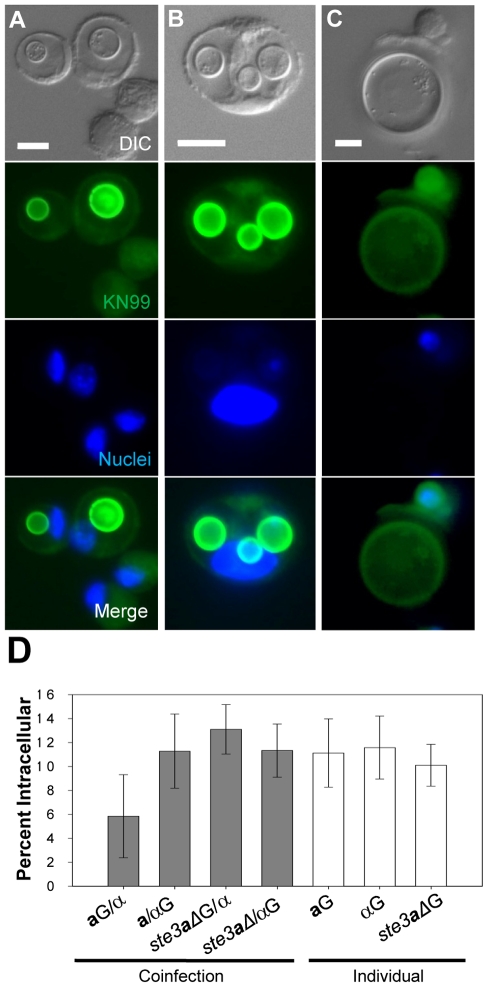
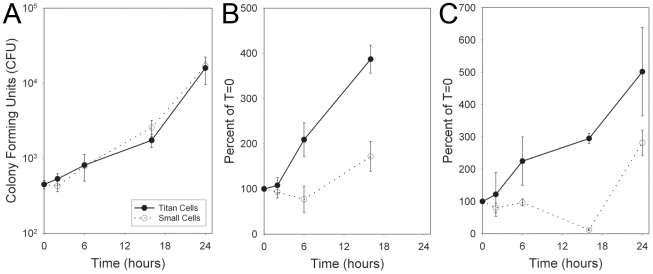
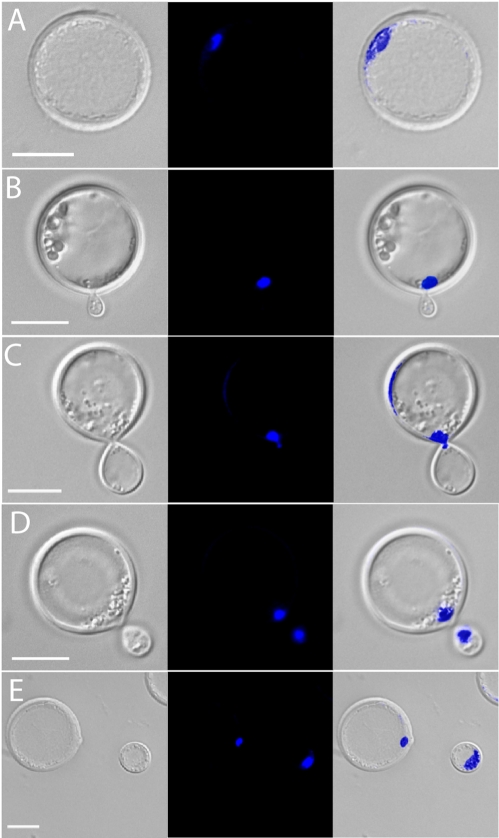
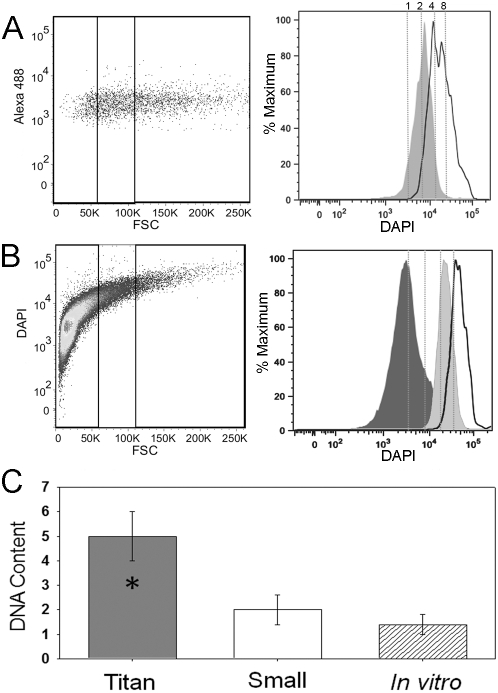
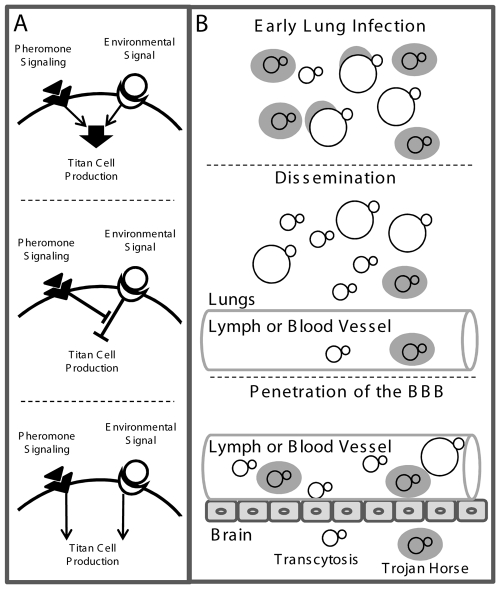
Cryptococcus neoformans is a common life-threatening human fungal pathogen. The size of cryptococcal cells is typically 5 to 10 microm. Cell enlargement was observed in vivo, producing cells up to 100 microm. These morphological changes in cell size affected pathogenicity via reducing phagocytosis by host mononuclear cells, increasing resistance to oxidative and nitrosative stress, and correlated with reduced penetration of the central nervous system. Cell enlargement was stimulated by coinfection with strains of opposite mating type, and ste3aDelta pheromone receptor mutant strains had reduced cell enlargement. Finally, analysis of DNA content in this novel cell type revealed that these enlarged cells were polyploid, uninucleate, and produced daughter cells in vivo. These results describe a novel mechanism by which C. neoformans evades host phagocytosis to allow survival of a subset of the population at early stages of infection. Thus, morphological changes play unique and specialized roles during infection.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- San-Blas G, Travassos LR, Fries BC, Goldman DL, Casadevall A, et al. Fungal morphogenesis and virulence. Med Mycol. 2000;38(Suppl 1):79–86. - PubMed
-
- Morrow CA, Fraser JA. Sexual reproduction and dimorphism in the pathogenic basidiomycetes. FEMS Yeast Res. 2009;9:161–177. - PubMed
-
- Hung CY, Xue J, Cole GT. Virulence mechanisms of Coccidioides. Ann N Y Acad Sci. 2007;1111:225–235. - PubMed
-
- Mavor AL, Thewes S, Hube B. Systemic fungal infections caused by Candida species: epidemiology, infection process and virulence attributes. Curr Drug Targets. 2005;6:863–874. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources