doi: 10.1128/JVI.00696-10.
Epub 2010 Jun 2.
Influence of pr-M cleavage on the heterogeneity of extracellular dengue virus particles
Affiliations
- PMID: 20519400
- PMCID: PMC2916513
- DOI: 10.1128/JVI.00696-10
Item in Clipboard
Influence of pr-M cleavage on the heterogeneity of extracellular dengue virus particles
Jiraphan Junjhon et al.
J Virol.
2010 Aug.
Display options
Format
Display options
Format
Abstract
During dengue virus replication, an incomplete cleavage of the envelope glycoprotein prM, generates a mixture of mature (prM-less) and prM-containing, immature extracellular particles. In this study, sequential immunoprecipitation and cryoelectron microscopy revealed a third type of extracellular particles, the partially mature particles, as the major prM-containing particles in a dengue serotype 2 virus. Changes in the proportion of viral particles in the pr-M junction mutants exhibiting altered levels of prM cleavage suggest that the partially mature particles may represent an intermediate subpopulation in the virus maturation pathway. These findings are consistent with a model suggesting the progressive mode of prM cleavage.
Figures
{kind=link}
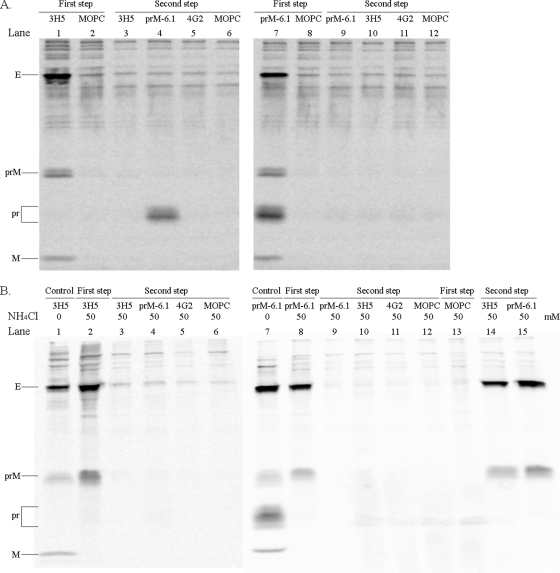
Sequential immunoprecipitation of extracellular viral particles. (A, left panel) The radiolabeled particles in a pretitrated volume (200 μl) of the culture fluid of virus-infected C6/36 cells were reacted with protein G Sepharose beads overnight and then with 3H5 (lane 1)- or MOPC-21 (lane 2)-coated beads in 1 ml at 4°C for 36 h. After a wash, particles were dissociated from beads with 2.5% 2-mercaptoethanol-1.5% SDS-40 mM Tris-HCl (pH 6.8)-0.02% bromophenol blue at 37°C for 30 min. In the coating step, 15 μg (3H5) or 10 μg (other antibodies) of protein G Sepharose column-purified antibodies and, in the majority of experiments, a fixed amount (5 μl) of an ascites, which was devoid of anti-dengue virus activity, were incubated with 30 μl of 50% (vol/vol) protein G Sepharose in 1 ml of 50 mM Tris-HCl (pH 7.4)-4 mM EDTA (pH 8.0)-300 mM NaCl overnight. Unbound particles from the 3H5 reaction were separated by spinning them in a microcentrifuge at 14,000 rpm for 5 min, and then precipitated with a set of beads that had been coated with 3H5 (lane 3), prM-6.1 (lane 4), 4G2 (lane 5), or MOPC-21 (lane 6). The eluted proteins were separated by electrophoresis in 0.1% SDS-15% polyacrylamide gel and the radioactivity signals captured with a phosphorimager. (A, right panel) The radiolabeled viral particles were reacted with protein G Sepharose and then with prM-6.1 (lane 7)- or MOPC-21 (lane 8)-coated beads overnight prior to elution. Subsequent precipitation of the unbound particles was performed with beads that had been coated with prM-6.1 (lane 9), 3H5 (lane 10), 4G2 (lane 11), or MOPC-21 (lane 12). (B) Viral particles derived from the 50 mM NH4Cl-treated, infected C6/36 culture were employed in the sequential immunoprecipitation with 3H5 (lanes 2 to 6) or prM-6.1 (lanes 8 to 12) as described for panel A. Lanes 1 and 7 represent the reaction of particles from NH4Cl-untreated cultures with 3H5 and prM-6.1, respectively, to ensure the correct use of intended antibodies. Lanes 13 to 15 represent an initial precipitation of particles from NH4Cl-treated cultures with MOPC-21 (lane 13) and subsequent reactions of unbound materials with 3H5 (lane 14) or prM-6.1 (lane 15). The antibody specificities were determined previously (15). The viral proteins are indicated. Note that C is not observed, as it is inefficiently labeled with the isotopes used.
{kind=link}
Cryo-EM visualization of extracellular virus particles. C6/36 cells were infected with dengue viruses at a multiplicity of infection of 0.05. After 5 to 7 days, the culture medium was collected and clarified by centrifugation at 6,000 rpm in a J2-21 rotor (Beckman) at 4°C for 30 min. Viral particles were precipitated with 8 g% polyethylene glycol (PEG) in 12 mM Tris (pH 8.0)-120 mM NaCl-1 mM EDTA buffer at 4°C for overnight. The precipitates were collected by spinning at 9,500 rpm for 50 min in the J2-21 rotor, resuspended with 12 mM Tris (pH 8.0)-120 mM NaCl-1 mM EDTA buffer, and then centrifuged in a 24 g% sucrose cushion (32,000 rpm for 1.5 h at 4°C, using a Beckman SW41 rotor). Particles were further purified in a 10- to 35-g% potassium tartrate gradient (32,000 rpm for 2 h at 4°C in a Beckman SW41 rotor). The visible viral band was collected, concentrated by using a centrifugal filter device, applied onto the carbon-coated cryoelectron grids, and vitrified in liquid ethane. The grids were examined with a transmission electron microscope. Images were recorded at a magnification of 45,000 under low-dose conditions (14 to 17 e−/Å2). The micrographs taken from each virus preparation were digitized at a 7-μm step size to yield a sampling of 2.96 Å/pixel. The morphology of virus particles was visualized using the RobEM program (http://cryoem.ucsd.edu/programDocs/runRobem.txt ). (A) The cryo-EM micrograph of the parent virus, strain 16681Nde(+). M, I, and P indicate the mature, immature, and partially mature particles, respectively. (B and C) Cropped cryo-EM images showing single particles of 16681Nde(+) and prH200A, respectively. The bar represents 100 nm.
{kind=link}
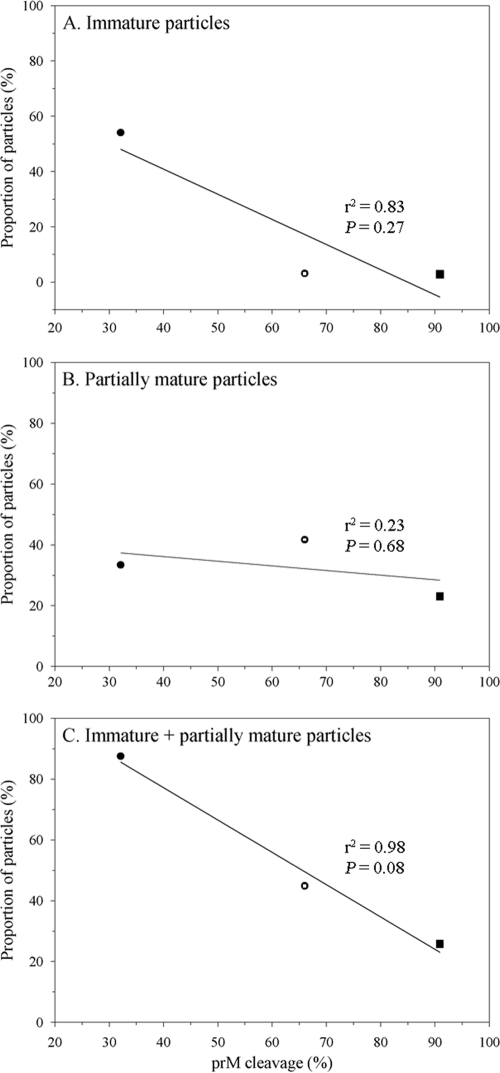
Relationship between the proportion of prM-containing extracellular particles and the extent of prM cleavage. The immature and partially mature particles present in the purified preparations of 16681Nde(+) (open circle), prH200A (closed circle), and prE203A (closed square) were categorized by cryo-EM based on their surface morphology, and the proportion of each particle subpopulation was plotted against the level of prM cleavage of each virus. The levels of prM cleavage of 16681Nde(+) and the pr-M junction mutants were determined in a previous study (15). Pearson's correlation and linear regression analysis were employed in the evaluation of the relationship between the level of prM cleavage and the proportion of particle subpopulations by using the STATA 8.2 software (StataCorp, TX).
References
-
- Blok, J., S. M. McWilliam, H. C. Butler, A. J. Gibbs, G. Weiller, B. L. Herring, A. C. Hemsley, J. G. Aaskov, S. Yoksan, and N. Bhamarapravati. 1992. Comparison of a dengue-2 virus and its candidate vaccine derivative: sequence relationships with the flaviviruses and other viruses. Virology 187:573-590. - PMC - PubMed
-
- Cherrier, M. V., B. Kaufmann, G. E. Nybakken, S.-M. Lok, J. T. Warren, B. R. Chen, C. A. Nelson, V. A. Kostyuchenko, H. A. Holdaway, P. R. Chipman, R. J. Kuhn, M. S. Diamond, M. G. Rossmann, and D. H. Fremont. 2009. Structural basis for the preferential recognition of immature flaviviruses by a fusion-loop antibody. EMBO J. 28:3269-3276. - PMC - PubMed
-
- Crabtree, M. B., R. C. Sang, V. Stollar, L. M. Dunster, and B. R. Miller. 2003. Genetic and phenotypic characterization of the newly described insect flavivirus, Kamiti River virus. Arch. Virol. 148:1095-1118. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials