Amiloride inhibits macropinocytosis by lowering submembranous pH and preventing Rac1 and Cdc42 signaling
- PMID: 20156964
- PMCID: PMC2828922
- DOI: 10.1083/jcb.200908086
Amiloride inhibits macropinocytosis by lowering submembranous pH and preventing Rac1 and Cdc42 signaling
Erratum in
- J Cell Biol. 2010 Apr 19;189(2):385
Abstract
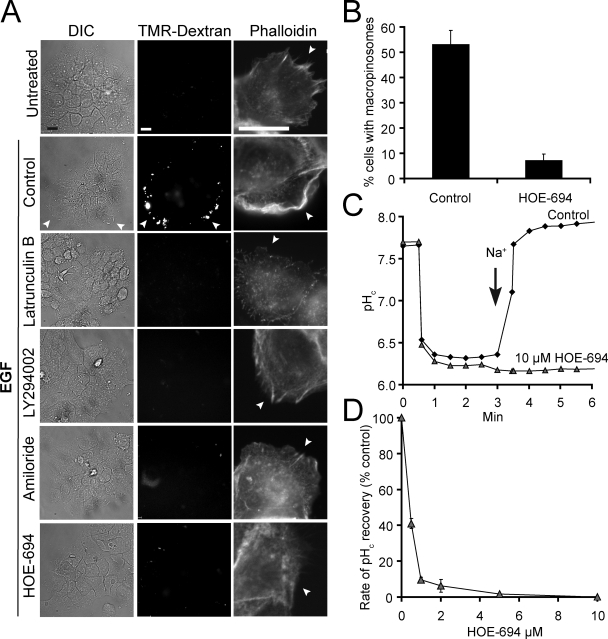
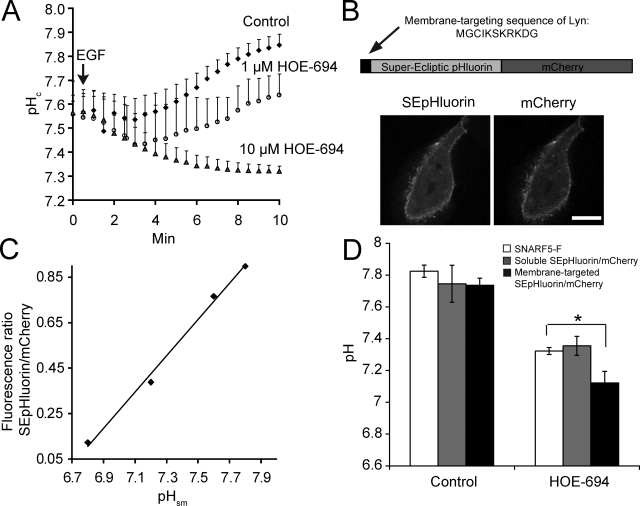
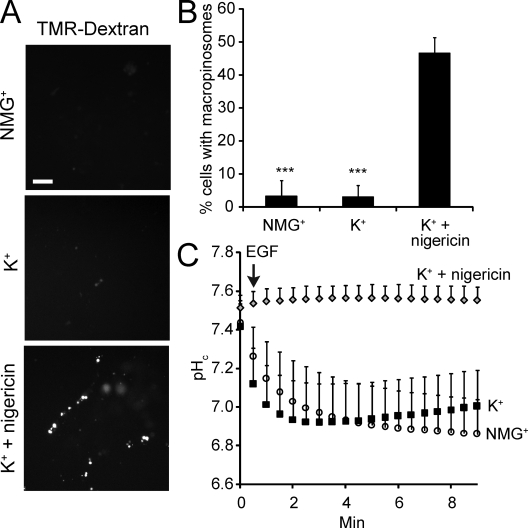
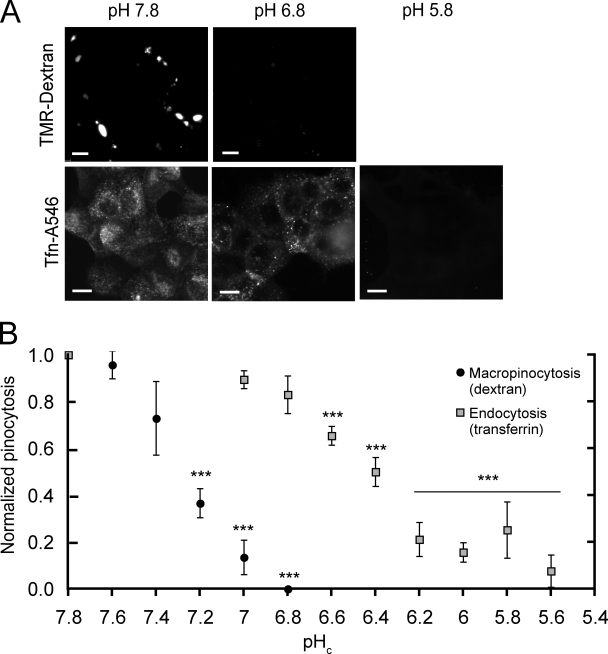
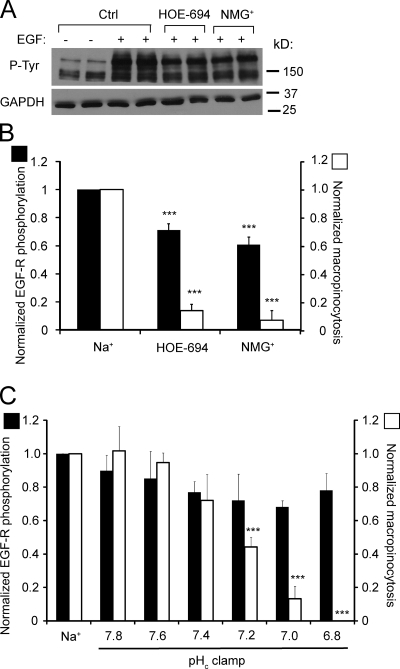
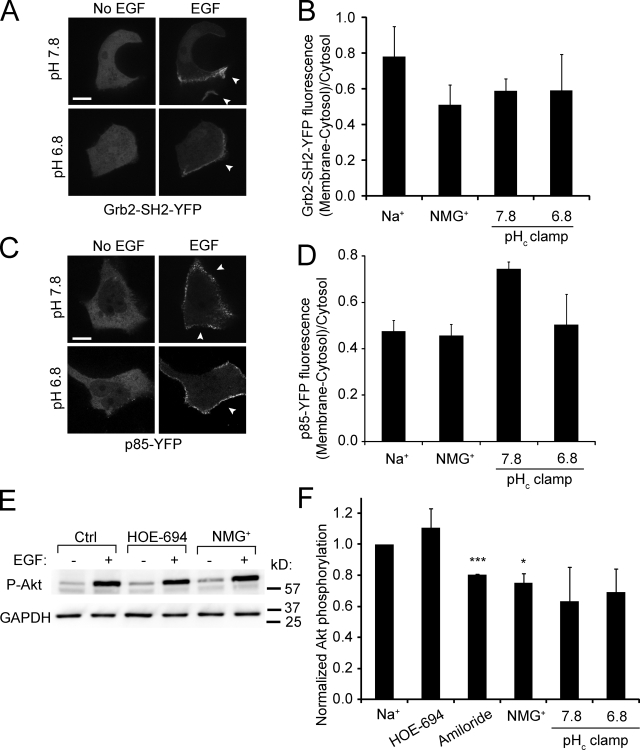
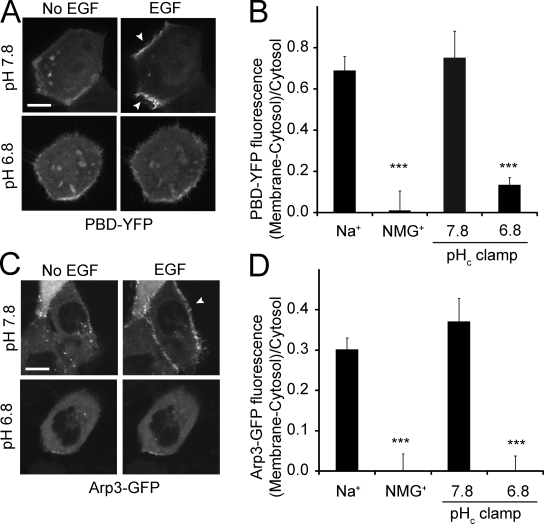
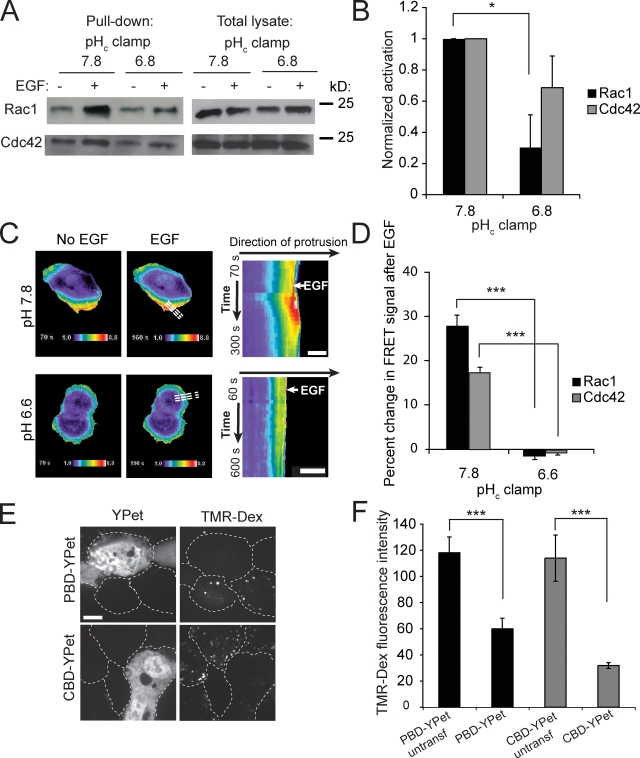
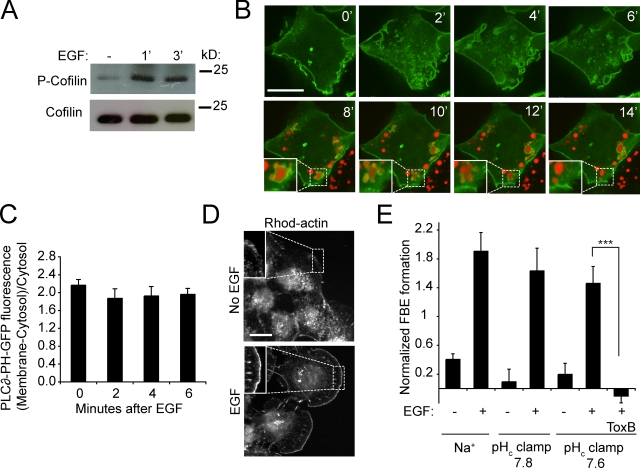
Macropinocytosis is differentiated from other types of endocytosis by its unique susceptibility to inhibitors of Na(+)/H(+) exchange. Yet, the functional relationship between Na(+)/H(+) exchange and macropinosome formation remains obscure. In A431 cells, stimulation by EGF simultaneously activated macropinocytosis and Na(+)/H(+) exchange, elevating cytosolic pH and stimulating Na(+) influx. Remarkably, although inhibition of Na(+)/H(+) exchange by amiloride or HOE-694 obliterated macropinocytosis, neither cytosolic alkalinization nor Na(+) influx were required. Instead, using novel probes of submembranous pH, we detected the accumulation of metabolically generated acid at sites of macropinocytosis, an effect counteracted by Na(+)/H(+) exchange and greatly magnified when amiloride or HOE-694 were present. The acidification observed in the presence of the inhibitors did not alter receptor engagement or phosphorylation, nor did it significantly depress phosphatidylinositol-3-kinase stimulation. However, activation of the GTPases that promote actin remodelling was found to be exquisitely sensitive to the submembranous pH. This sensitivity confers to macropinocytosis its unique susceptibility to inhibitors of Na(+)/H(+) exchange.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous