C-terminal mutants of apolipoprotein L-I efficiently kill both Trypanosoma brucei brucei and Trypanosoma brucei rhodesiense
- PMID: 19997494
- PMCID: PMC2778949
- DOI: 10.1371/journal.ppat.1000685
C-terminal mutants of apolipoprotein L-I efficiently kill both Trypanosoma brucei brucei and Trypanosoma brucei rhodesiense
Abstract
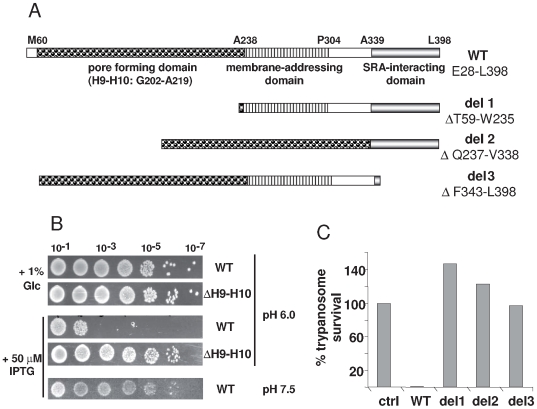
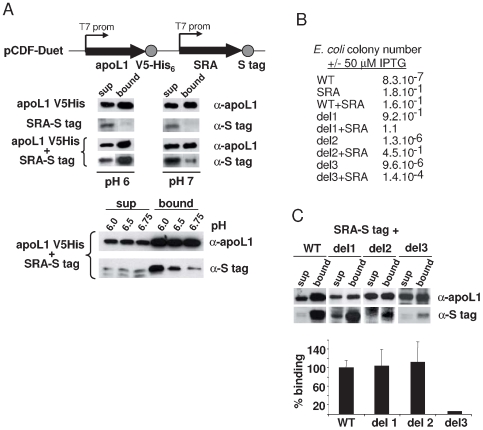
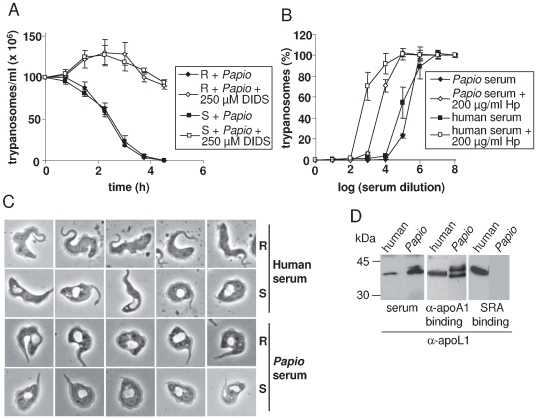
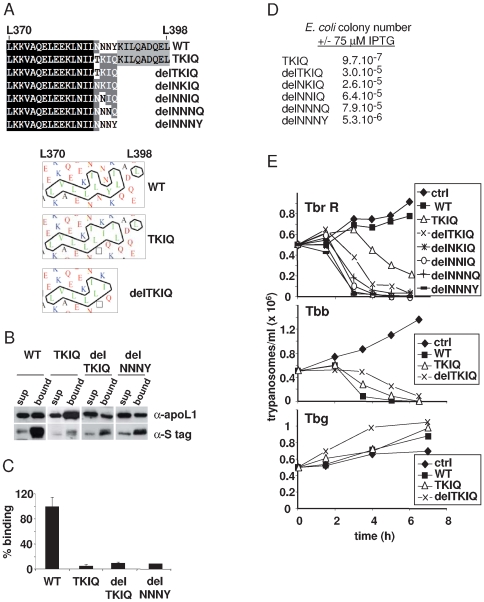
Apolipoprotein L-I (apoL1) is a human-specific serum protein that kills Trypanosoma brucei through ionic pore formation in endosomal membranes of the parasite. The T. brucei subspecies rhodesiense and gambiense resist this lytic activity and can infect humans, causing sleeping sickness. In the case of T. b. rhodesiense, resistance to lysis involves interaction of the Serum Resistance-Associated (SRA) protein with the C-terminal helix of apoL1. We undertook a mutational and deletional analysis of the C-terminal helix of apoL1 to investigate the linkage between interaction with SRA and lytic potential for different T. brucei subspecies. We confirm that the C-terminal helix is the SRA-interacting domain. Although in E. coli this domain was dispensable for ionic pore-forming activity, its interaction with SRA resulted in inhibition of this activity. Different mutations affecting the C-terminal helix reduced the interaction of apoL1 with SRA. However, mutants in the L370-L392 leucine zipper also lost in vitro trypanolytic activity. Truncating and/or mutating the C-terminal sequence of human apoL1 like that of apoL1-like sequences of Papio anubis resulted in both loss of interaction with SRA and acquired ability to efficiently kill human serum-resistant T. b. rhodesiense parasites, in vitro as well as in transgenic mice. These findings demonstrate that SRA interaction with the C-terminal helix of apoL1 inhibits its pore-forming activity and determines resistance of T. b. rhodesiense to human serum. In addition, they provide a possible explanation for the ability of Papio serum to kill T. b. rhodesiense, and offer a perspective to generate transgenic cattle resistant to both T. b. brucei and T. b. rhodesiense.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Vanhamme L, Paturiaux-Hanocq F, Poelvoorde P, Nolan DP, Lins L, et al. Apolipoprotein L-I is the trypanosome lytic factor of human serum. Nature. 2003;422:83–87. - PubMed
-
- Pays E, Vanhollebeke B, Vanhamme L, Paturiaux-Hanocq F, Nolan DP, et al. The trypanolytic factor of human serum. Nat Rev Microbiol. 2006;4:477–486. - PubMed
-
- Vanhollebeke B, Demuylder G, Nielsen MJ, Pays A, Tebabi P, et al. A haptoglobin-hemoglobin receptor conveys innate immunity to Trypanosoma brucei in humans. Science. 2008;320:677–681. - PubMed
-
- Pérez-Morga D, Vanhollebeke B, Paturiaux-Hanocq F, Nolan DP, Lins L, et al. Apolipoprotein L-I promotes trypanosome lysis by forming pores in lysosomal membranes. Science. 2005;309:469–472. - PubMed
-
- Xong HV, Vanhamme L, Chamekh M, Chimfwembe CE, Van den Abbeele J, et al. A VSG expression site-associated gene confers resistance to human serum in Trypanosoma rhodesiense. Cell. 1998;95:839–846. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous