Role of the Plasmodium export element in trafficking parasite proteins to the infected erythrocyte
- PMID: 19055692
- PMCID: PMC2682620
- DOI: 10.1111/j.1600-0854.2008.00864.x
Role of the Plasmodium export element in trafficking parasite proteins to the infected erythrocyte
Abstract
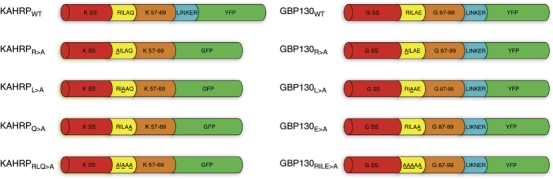
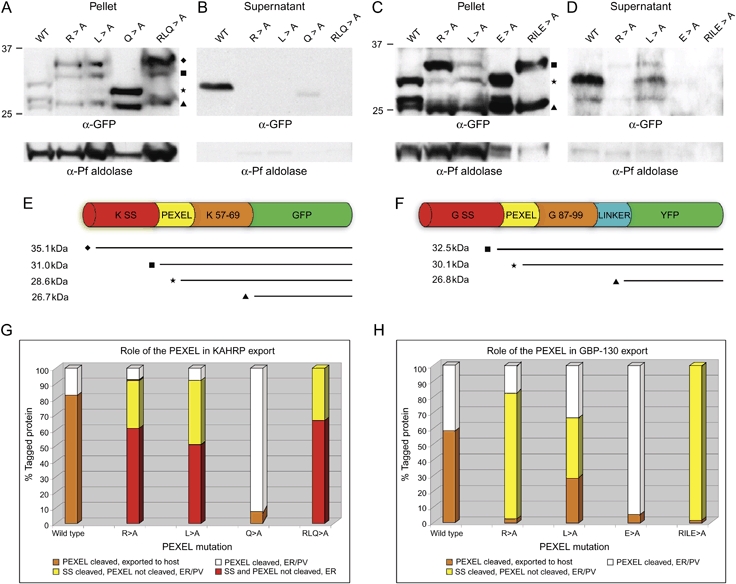
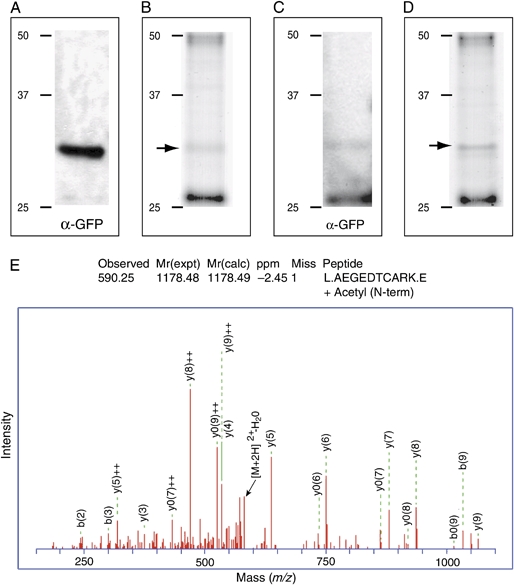
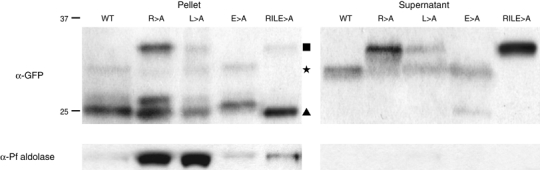
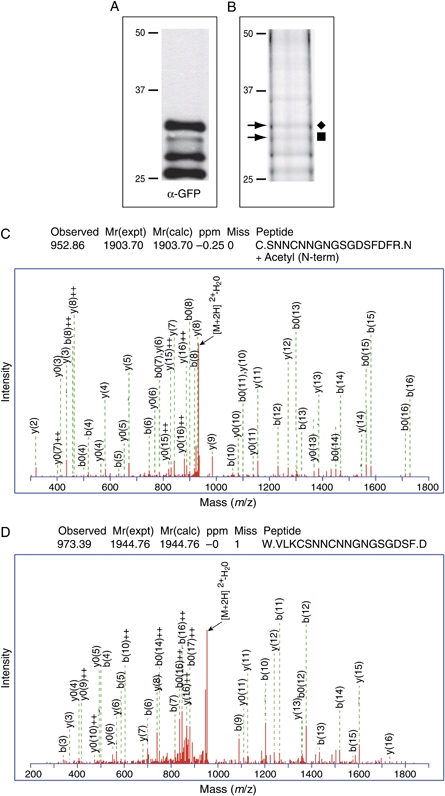
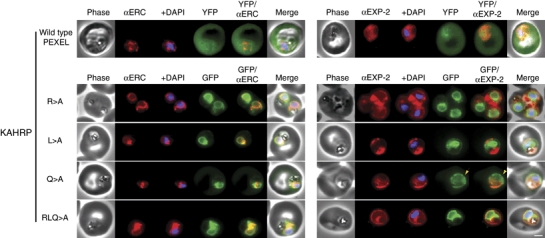
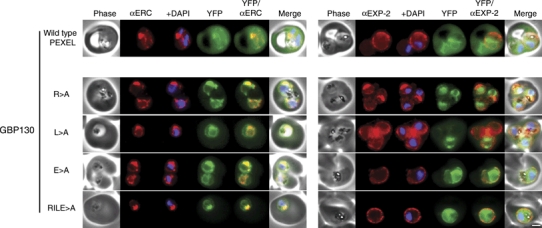
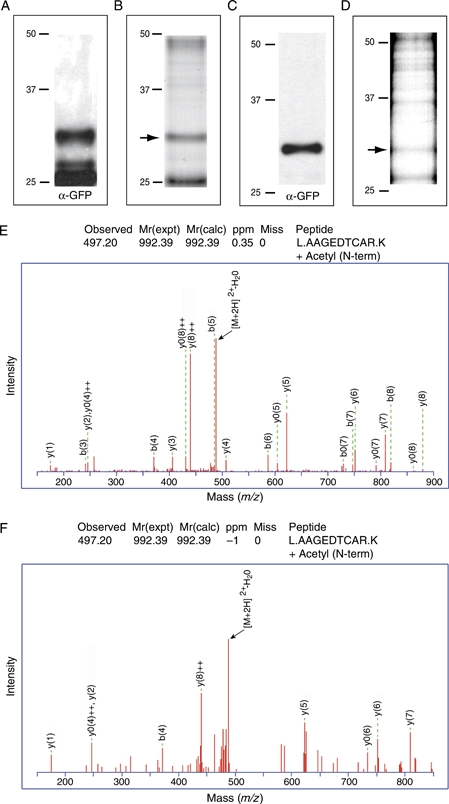
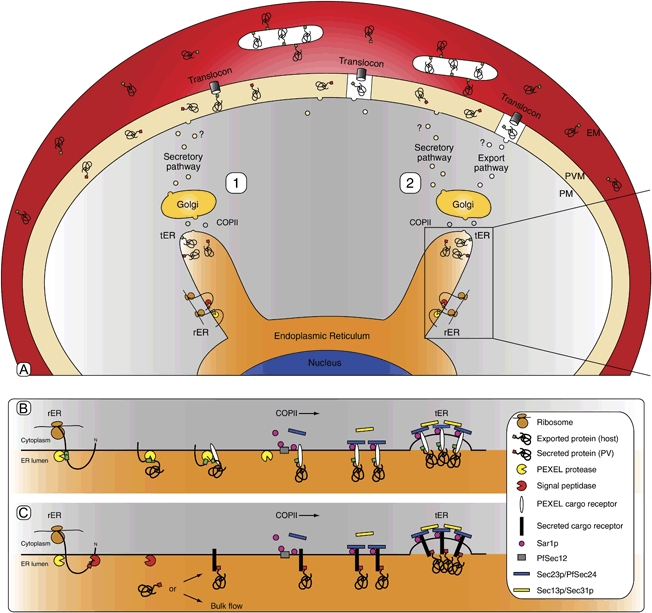
The intracellular survival of Plasmodium falciparum within human erythrocytes is dependent on export of parasite proteins that remodel the host cell. Most exported proteins require a conserved motif (RxLxE/Q/D), termed the Plasmodium export element (PEXEL) or vacuolar targeting sequence (VTS), for targeting beyond the parasitophorous vacuole membrane and into the host cell; however, the precise role of this motif in export is poorly defined. We used transgenic P. falciparum expressing chimeric proteins to investigate the function of the PEXEL motif for export. The PEXEL constitutes a bifunctional export motif comprising a protease recognition sequence that is cleaved, in the endoplasmic reticulum, from proteins destined for export, in a PEXEL arginine- and leucine-dependent manner. Following processing, the remaining conserved PEXEL residue is required to direct the mature protein to the host cell. Furthermore, we demonstrate that N acetylation of proteins following N-terminal processing is a PEXEL-independent process that is insufficient for correct export to the host cell. This work defines the role of each residue in the PEXEL for export into the P. falciparum-infected erythrocyte.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Baruch DI, Pasloske BL, Singh HB, Bi X, Ma XC, Feldman M, Taraschi TF, Howard RJ. Cloning the P.falciparum gene encoding PfEMP1, a malarial variant antigen and adherence receptor on the surface of parasitized human erythrocytes. Cell. 1995;82:77–87. - PubMed
-
- Su X-Z, Kirkman LA, Fujioka H, Wellems TE. Complex polymorphisms in an ∼330 kDa protein are linked to chloroquine-resistant P. falciparum in Southeast Asia and Africa. Cell. 1997;91:593–603. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources