The neglected role of antibody in protection against bacteremia caused by nontyphoidal strains of Salmonella in African children
- PMID: 18357343
- PMCID: PMC2268878
- DOI: 10.1172/JCI33998
The neglected role of antibody in protection against bacteremia caused by nontyphoidal strains of Salmonella in African children
Abstract
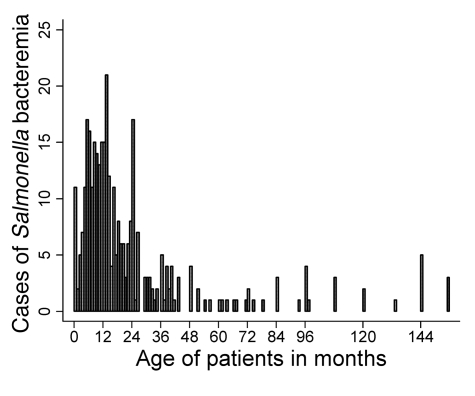
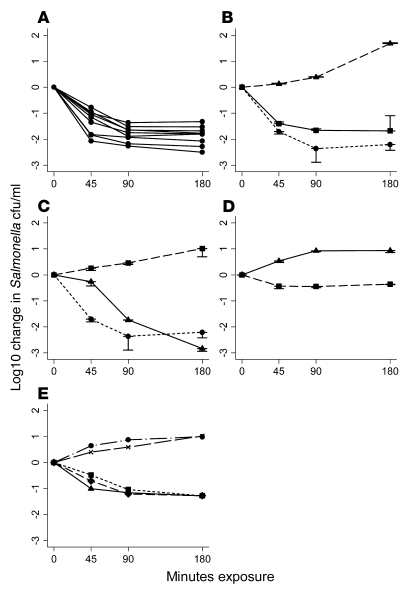
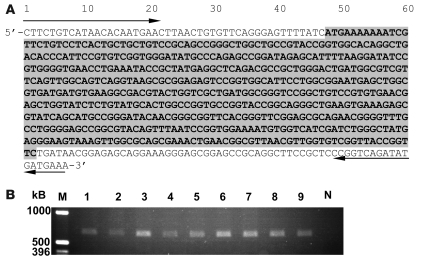
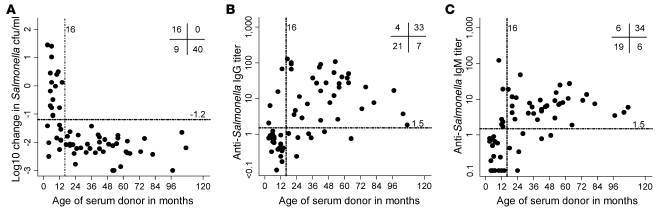
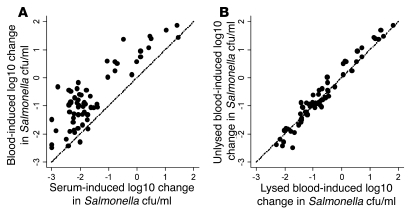
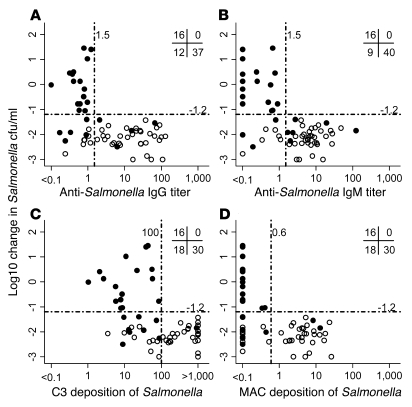
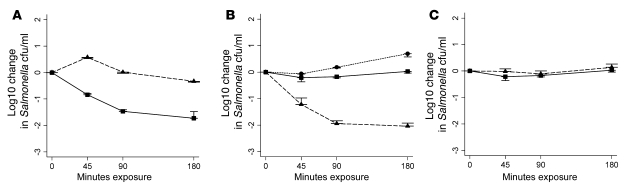
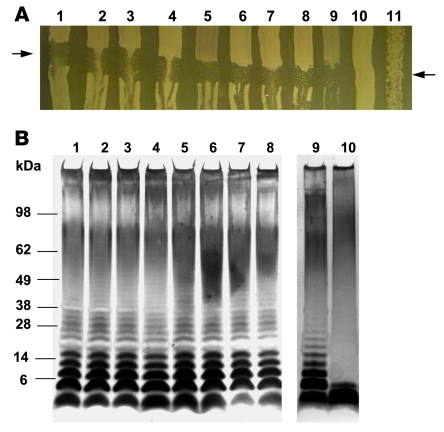
Nontyphoidal strains of Salmonella (NTS) are a common cause of bacteremia among African children. Cell-mediated immune responses control intracellular infection, but they do not protect against extracellular growth of NTS in the blood. We investigated whether antibody protects against NTS bacteremia in Malawian children, because we found this condition mainly occurs before 2 years of age, with relative sparing of infants younger than 4 months old. Sera from all healthy Malawian children tested aged more than 16 months contained anti-Salmonella antibody and successfully killed NTS. Killing was mediated by complement membrane attack complex and not augmented in the presence of blood leukocytes. Sera from most healthy children less than 16 months old lacked NTS-specific antibody, and sera lacking antibody did not kill NTS despite normal complement function. Addition of Salmonella-specific antibody, but not mannose-binding lectin, enabled NTS killing. All NTS strains tested had long-chain lipopolysaccharide and the rck gene, features that resist direct complement-mediated killing. Disruption of lipopolysaccharide biosynthesis enabled killing of NTS by serum lacking Salmonella-specific antibody. We conclude that Salmonella-specific antibody that overcomes the complement resistance of NTS develops by 2 years of life in Malawian children. This finding and the age-incidence of NTS bacteremia suggest that antibody protects against NTS bacteremia and support the development of vaccines against NTS that induce protective antibody.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Graham S.M. Salmonellosis in children in developing and developed countries and populations. Curr. Opin. Infect. Dis. 2002;15:507–512. - PubMed