Adaptations of Trypanosoma brucei to gradual loss of kinetoplast DNA: Trypanosoma equiperdum and Trypanosoma evansi are petite mutants of T. brucei
- PMID: 18245376
- PMCID: PMC2538871
- DOI: 10.1073/pnas.0711799105
Adaptations of Trypanosoma brucei to gradual loss of kinetoplast DNA: Trypanosoma equiperdum and Trypanosoma evansi are petite mutants of T. brucei
Abstract
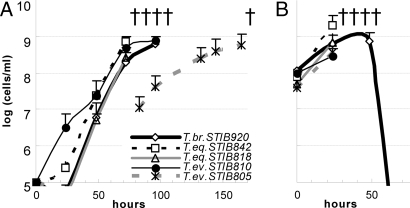
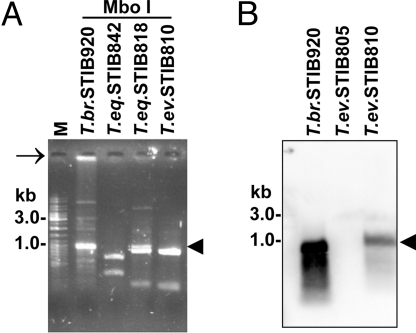
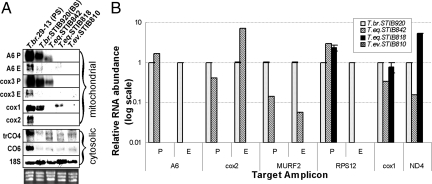
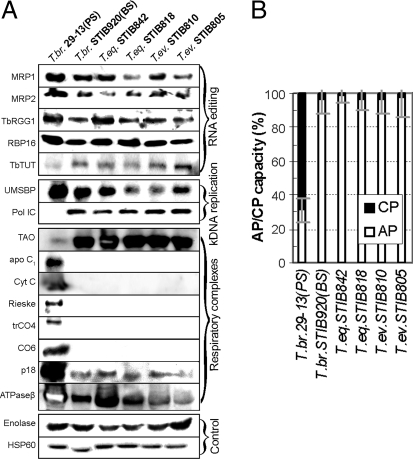
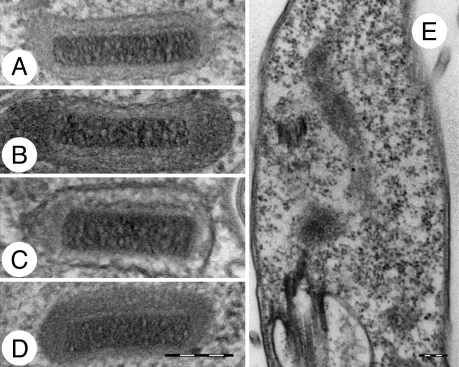
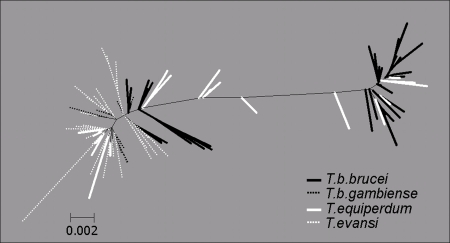
Trypanosoma brucei is a kinetoplastid flagellate, the agent of human sleeping sickness and ruminant nagana in Africa. Kinetoplastid flagellates contain their eponym kinetoplast DNA (kDNA), consisting of two types of interlocked circular DNA molecules: scores of maxicircles and thousands of minicircles. Maxicircles have typical mitochondrial genes, most of which are translatable only after RNA editing. Minicircles encode guide RNAs, required for decrypting the maxicircle transcripts. The life cycle of T. brucei involves a bloodstream stage (BS) in vertebrates and a procyclic stage (PS) in the tsetse fly vector. Partial [dyskinetoplastidy (Dk)] or total [akinetoplastidy (Ak)] loss of kDNA locks the trypanosome in the BS form. Transmission between vertebrates becomes mechanical without PS and tsetse mediation, allowing the parasite to spread outside the African tsetse belt. Trypanosoma equiperdum and Trypanosoma evansi are agents of dourine and surra, diseases of horses, camels, and water buffaloes. We have characterized representative strains of T. equiperdum and T. evansi by numerous molecular and classical parasitological approaches. We show that both species are actually strains of T. brucei, which lost part (Dk) or all (Ak) of their kDNA. These trypanosomes are not monophyletic clades and do not qualify for species status. They should be considered two subspecies, respectively T. brucei equiperdum and T. brucei evansi, which spontaneously arose recently. Dk/Ak trypanosomes may potentially emerge repeatedly from T. brucei.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Simpson AGB, Stevens JR, Lukeš J. The evolution and diversity of kinetoplastid flagellates. Trends Parasitol. 2006;22:168–174. - PubMed
-
- Lukeš J, Hashimi H, Zíková A. Unexplained complexity of the mitochondrial genome and transcriptome in kinetoplastid flagellates. Curr Genet. 2005;48:277–299. - PubMed
-
- Stuart KD, Schnaufer A, Ernst NL, Panigrahi AK. Complex management: RNA editing in trypanosomes. Trends Biochem Sci. 2005;30:97–105. - PubMed
-
- Shlomai J. The structure and replication of kinetoplast DNA. Curr Mol Med. 2004;4:623–647. - PubMed
-
- Liu BY, Liu YN, Motyka SA, Agbo EEC, Englund PT. Fellowship of the rings: the replication of kinetoplast DNA. Trends Parasitol. 2005;21:363–369. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources