Apical maxi-K (KCa1.1) channels mediate K+ secretion by the mouse submandibular exocrine gland
- PMID: 18216162
- PMCID: PMC3298180
- DOI: 10.1152/ajpcell.00511.2007
Apical maxi-K (KCa1.1) channels mediate K+ secretion by the mouse submandibular exocrine gland
Abstract
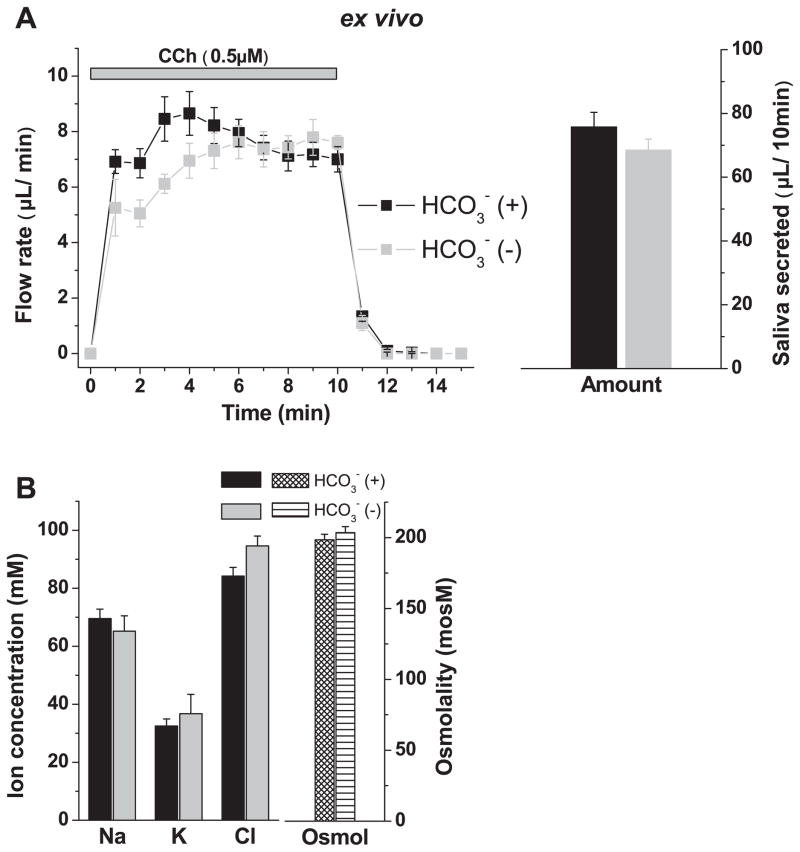
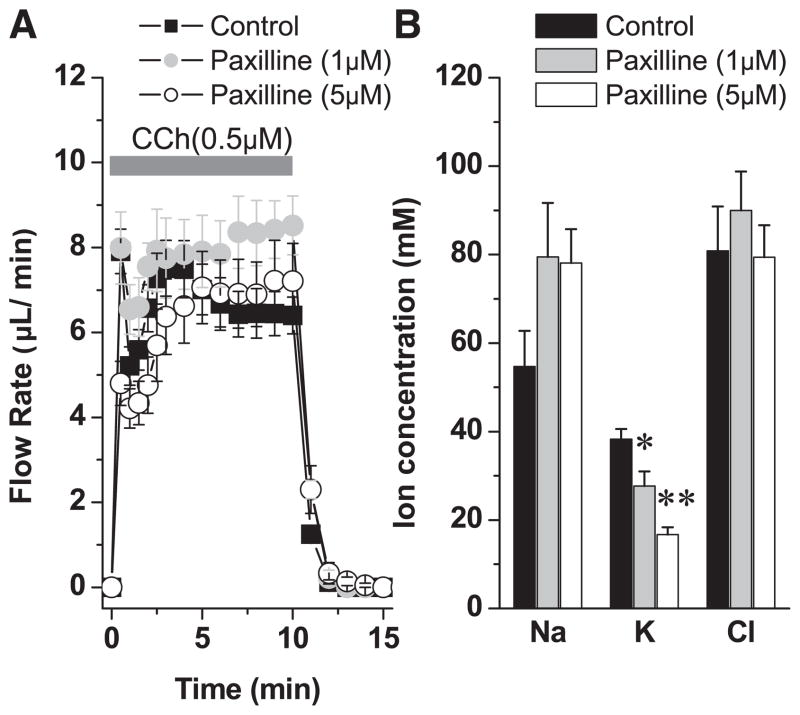
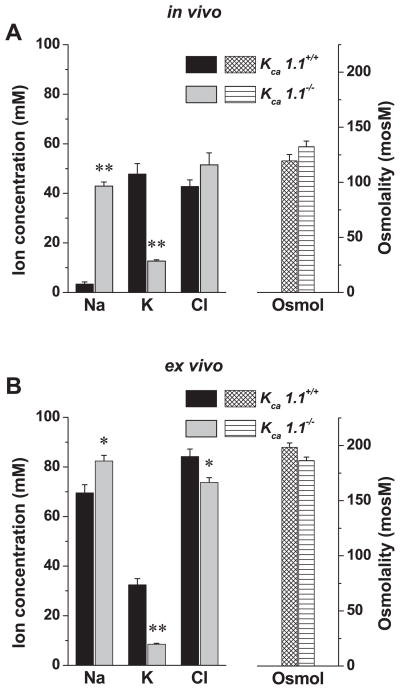
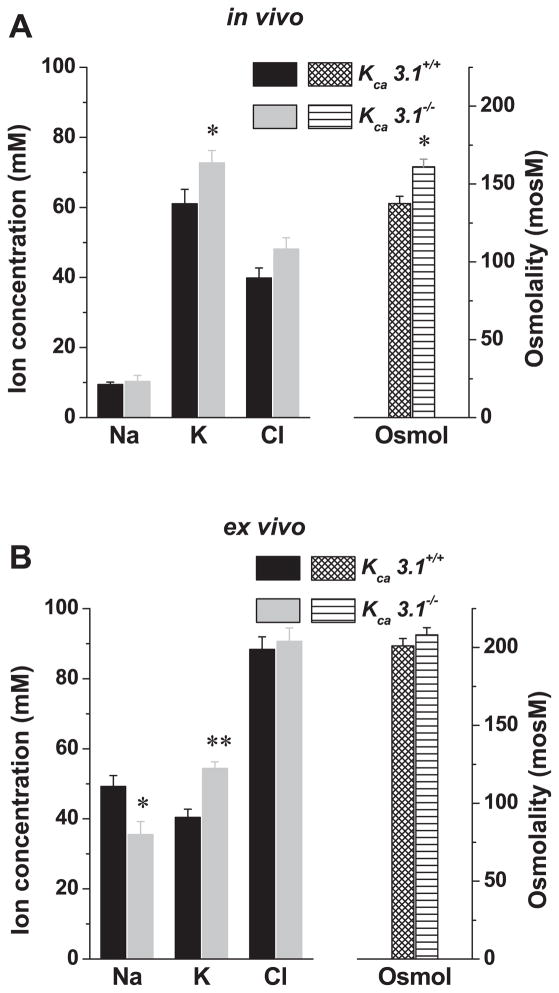
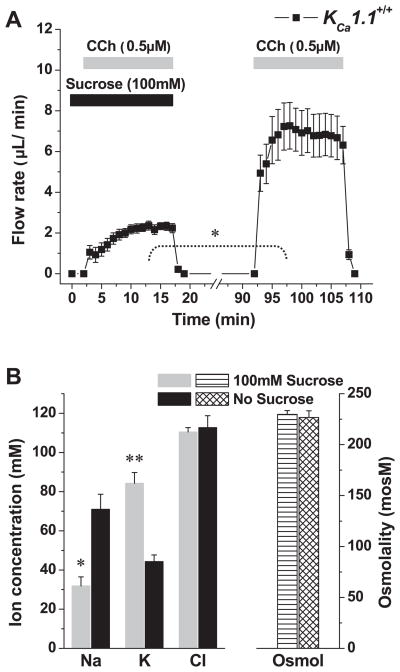
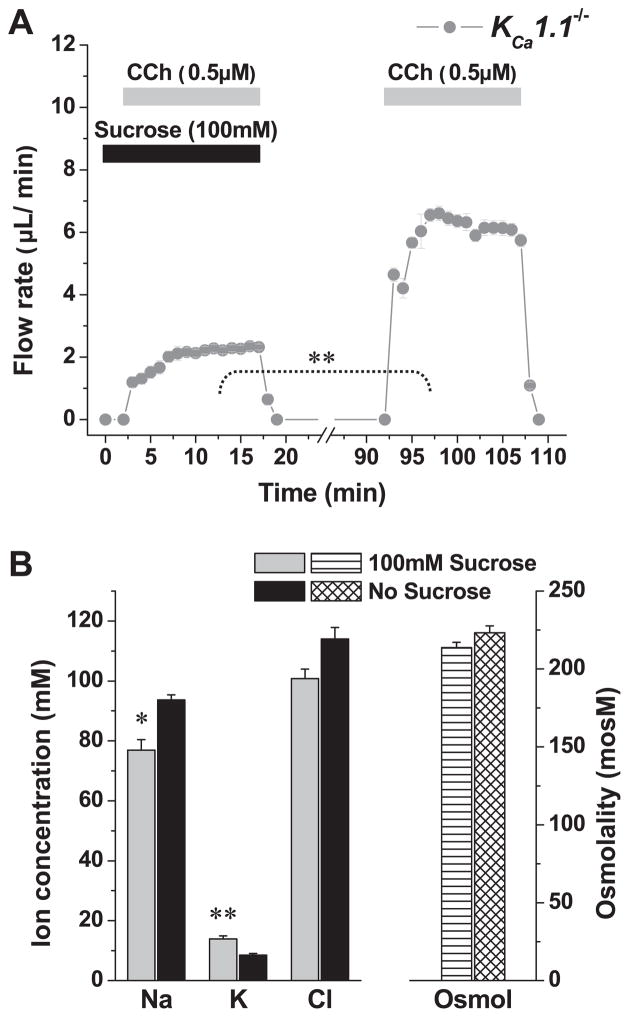
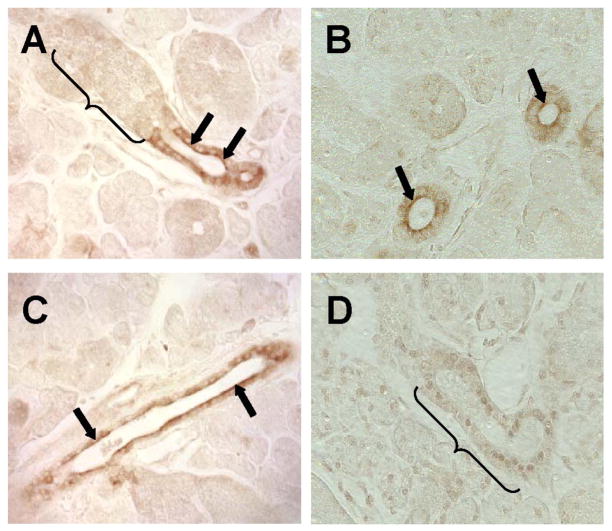
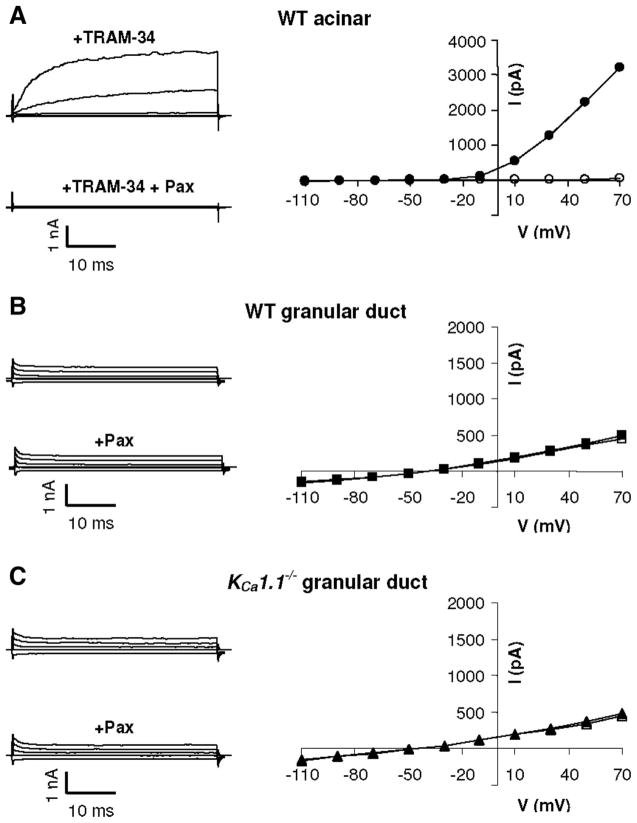
The exocrine salivary glands of mammals secrete K+ by an unknown pathway that has been associated with HCO3(-) efflux. However, the present studies found that K+ secretion in the mouse submandibular gland did not require HCO3(-), demonstrating that neither K+/HCO3(-) cotransport nor K+/H+ exchange mechanisms were involved. Because HCO3(-) did not appear to participate in this process, we tested whether a K channel is required. Indeed, K+ secretion was inhibited >75% in mice with a null mutation in the maxi-K, Ca2+-activated K channel (KCa1.1) but was unchanged in mice lacking the intermediate-conductance IKCa1 channel (KCa3.1). Moreover, paxilline, a specific maxi-K channel blocker, dramatically reduced the K+ concentration in submandibular saliva. The K+ concentration of saliva is well known to be flow rate dependent, the K+ concentration increasing as the flow decreases. The flow rate dependence of K+ secretion was nearly eliminated in KCa1.1 null mice, suggesting an important role for KCa1.1 channels in this process as well. Importantly, a maxi-K-like current had not been previously detected in duct cells, the theoretical site of K+ secretion, but we found that KCa1.1 channels localized to the apical membranes of both striated and excretory duct cells, but not granular duct cells, using immunohistochemistry. Consistent with this latter observation, maxi-K currents were not detected in granular duct cells. Taken together, these results demonstrate that the secretion of K+ requires and is likely mediated by KCa1.1 potassium channels localized to the apical membranes of striated and excretory duct cells in the mouse submandibular exocrine gland.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Alexander JH, van Lennep EW, Young JA. Water and electrolyte secretion by the exorbital lacrimal gland of the rat studied by micropuncture and catheterization techniques. Pflügers Arch. 1972;337:299–309. - PubMed
-
- Begenisich T, Nakamoto T, Ovitt CE, Nehrke K, Brugnara C, Alper SL, Melvin JE. Physiological roles of the intermediate conductance, Ca2+-activated potassium channel Kcnn4. J Biol Chem. 2004;279:47681–47687. - PubMed
-
- Botelho SY, Martinez EV. Electrolytes in lacrimal gland fluid and in tears at various flow rates in the rabbit. Am J Physiol. 1973;225:606–609. - PubMed
-
- Cook DI, Van Lennep EW, Roberts ML, Young JA. Secretion by the major salivary glands. In: Johnson LR, editor. Physiology of the Gastrointestinal Tract. 3. New York: Raven Press; 1994. pp. 1061–1117.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous