Molecular sabotage of plant defense by aphid saliva
- PMID: 17553961
- PMCID: PMC1965548
- DOI: 10.1073/pnas.0703535104
Molecular sabotage of plant defense by aphid saliva
Abstract
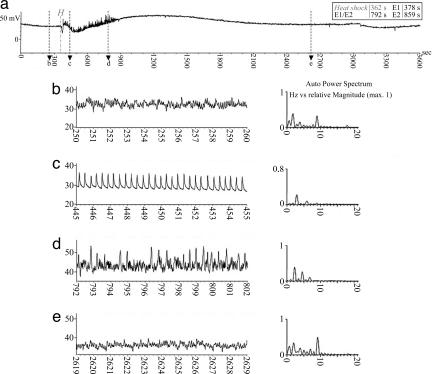
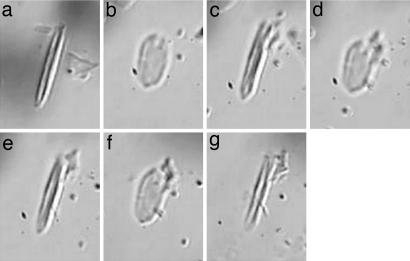
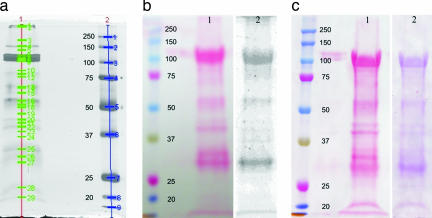
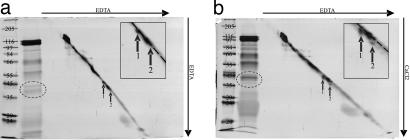
Aphids, which constitute one of the most important groups of agricultural pests, ingest nutrients from sieve tubes, the photoassimilate transport conduits in plants. Aphids are able to successfully puncture sieve tubes with their piercing mouthparts (stylets) and ingest phloem sap without eliciting the sieve tubes' normal occlusion response to injury. Occlusion mechanisms are calcium-triggered and may be prevented by chemical constituents in aphid saliva injected into sieve tubes before and during feeding. We recorded aphid feeding behavior with the electrical penetration graph (EPG) technique and then experimentally induced sieve tube plugging. Initiation of sieve tube occlusion caused a change in aphid behavior from phloem sap ingestion to secretion of watery saliva. Direct proof of "unplugging" properties of aphid saliva was provided by the effect of aphid saliva on forisomes. Forisomes are proteinaceous inclusions in sieve tubes of legumes that show calcium-regulated changes in conformation between a contracted state (below calcium threshold) that does not occlude the sieve tubes and a dispersed state (above calcium threshold) that occludes the sieve tubes. We demonstrated in vitro that aphid saliva induces dispersed forisomes to revert back to the nonplugging contracted state. Labeling Western-blotted saliva proteins with 45Ca2+ or ruthenium red inferred the presence of calcium-binding domains. These results demonstrate that aphid saliva has the ability to prevent sieve tube plugging by molecular interactions between salivary proteins and calcium. This provides aphids with access to a continuous flow of phloem sap and is a critical adaptation instrumental in the evolutionary success of aphids.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- van Bel AJE. Plant Cell Environ. 2003;26:125–149.
-
- van Bel AJE. Baluska F, Volkmann D, Barlow PW, editors. Cell–Cell Channels. 2006:1–6. www.Eurekah.com.
-
- Knoblauch M, van Bel AJE. Plant Cell. 1998;10:35–50.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous