Formation of hepatitis B virus covalently closed circular DNA: removal of genome-linked protein
- PMID: 17409153
- PMCID: PMC1900077
- DOI: 10.1128/JVI.02721-06
Formation of hepatitis B virus covalently closed circular DNA: removal of genome-linked protein
Abstract
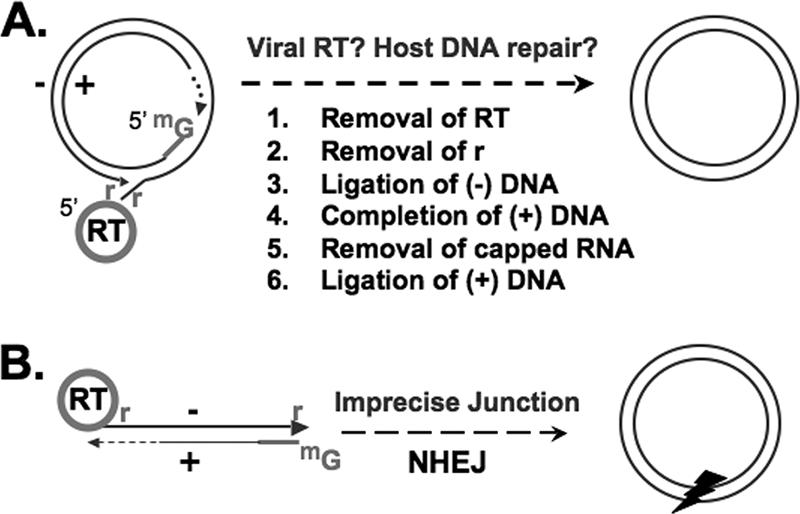
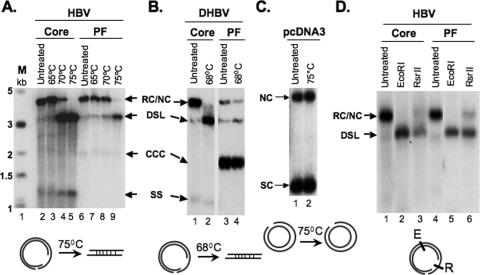
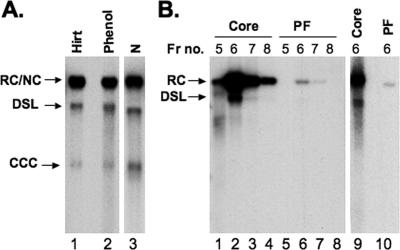
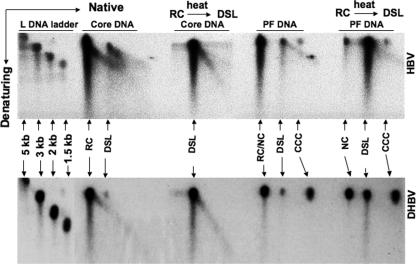
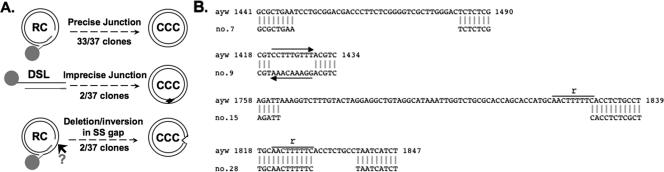
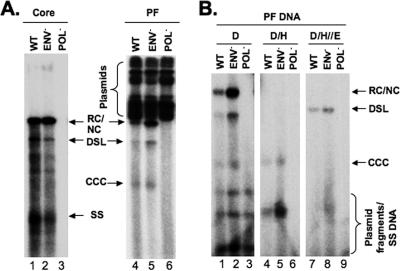
Hepatitis B virus (HBV) contains a small, partially double-stranded, relaxed circular (RC) DNA genome. RC DNA needs to be converted to covalently closed circular (CCC) DNA, which serves as the template for all viral RNA transcription. As a first step toward understanding how CCC DNA is formed, we analyzed the viral and host factors that may be involved in CCC DNA formation, using transient and stable DNA transfections of HBV and the related avian hepadnavirus, duck hepatitis B virus (DHBV). Our results show that HBV CCC DNA formed in hepatoma cells was derived predominantly from RC DNA with a precise junction sequence. In contrast to that of DHBV, HBV CCC DNA formation in cultured cells was accompanied by the accumulation of a RC DNA species from which the covalently attached viral reverse transcriptase (RT) protein was removed (protein-free or PF-RC DNA). Furthermore, whereas envelope deficiency led to increased CCC DNA formation in DHBV, it resulted mainly in increased PF-RC, but not CCC, DNA in HBV, suggesting that the envelope protein(s) may negatively regulate a step in CCC DNA formation that precedes deproteination in both HBV and DHBV. Interestingly, PF-RC DNA, in contrast to RT-linked RC DNA, contained, almost exclusively, mature plus-strand DNA, suggesting that the RT protein was removed preferentially from mature RC DNA.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Anderson, A. L., K. E. Banks, M. Pontoglio, M. Yaniv, and A. McLachlan. 2005. Alpha/beta interferon differentially modulates the clearance of cytoplasmic encapsidated replication intermediates and nuclear covalently closed circular hepatitis B virus (HBV) DNA from the livers of hepatocyte nuclear factor 1α-null HBV transgenic mice. J. Virol. 79:11045-11052. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials