The role of zooxanthellae in the thermal tolerance of corals: a 'nugget of hope' for coral reefs in an era of climate change
- PMID: 16928632
- PMCID: PMC1636081
- DOI: 10.1098/rspb.2006.3567
The role of zooxanthellae in the thermal tolerance of corals: a 'nugget of hope' for coral reefs in an era of climate change
Abstract
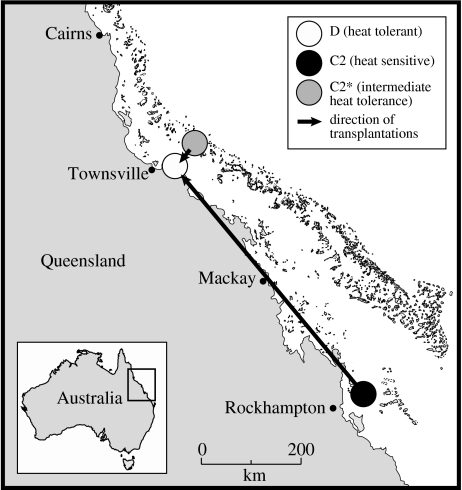
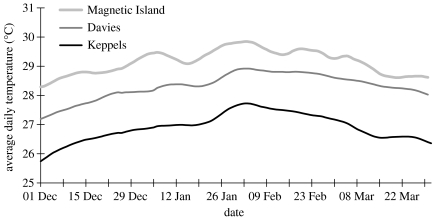
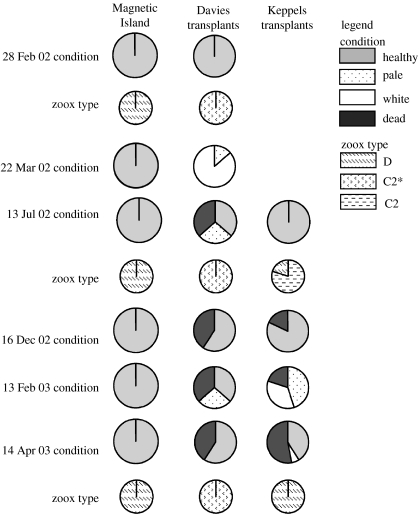
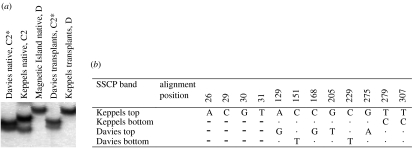
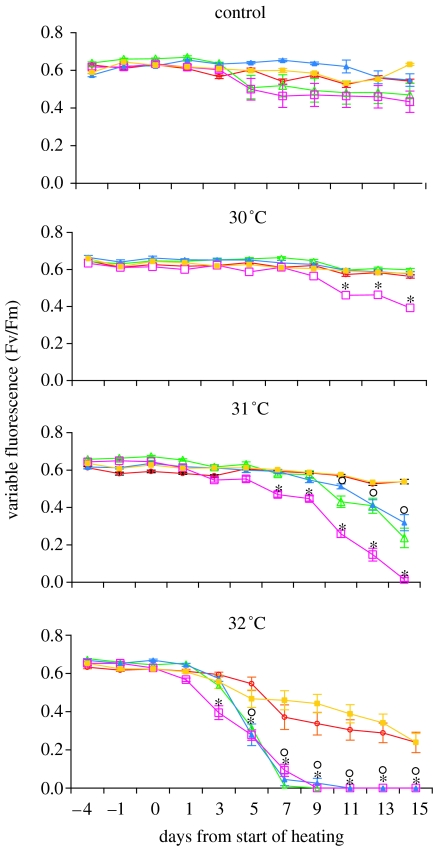
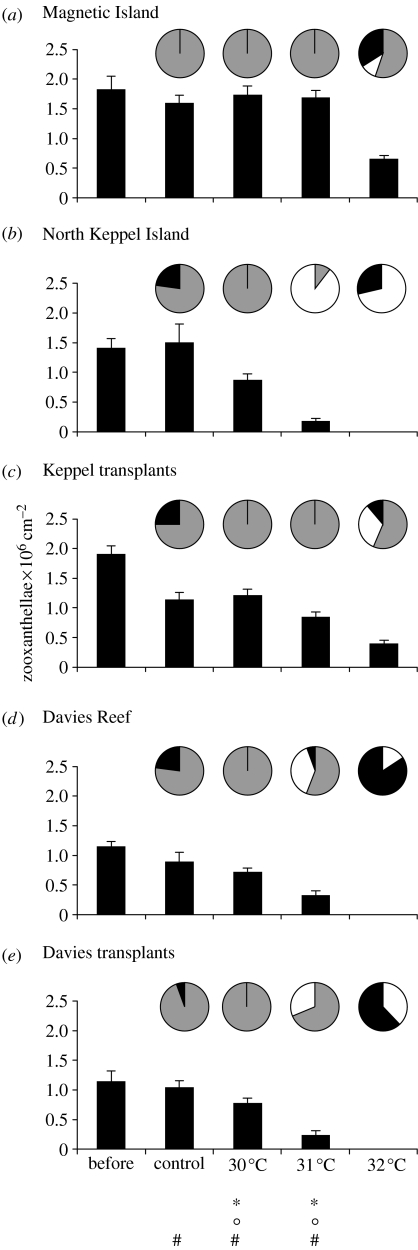
The ability of coral reefs to survive the projected increases in temperature due to global warming will depend largely on the ability of corals to adapt or acclimatize to increased temperature extremes over the next few decades. Many coral species are highly sensitive to temperature stress and the number of stress (bleaching) episodes has increased in recent decades. We investigated the acclimatization potential of Acropora millepora, a common and widespread Indo-Pacific hard coral species, through transplantation and experimental manipulation. We show that adult corals, at least in some circumstances, are capable of acquiring increased thermal tolerance and that the increased tolerance is a direct result of a change in the symbiont type dominating their tissues from Symbiodinium type C to D. Our data suggest that the change in symbiont type in our experiment was due to a shuffling of existing types already present in coral tissues, not through exogenous uptake from the environment. The level of increased tolerance gained by the corals changing their dominant symbiont type to D (the most thermally resistant type known) is around 1-1.5 degrees C. This is the first study to show that thermal acclimatization is causally related to symbiont type and provides new insight into the ecological advantage of corals harbouring mixed algal populations. While this increase is of huge ecological significance for many coral species, in the absence of other mechanisms of thermal acclimatization/adaptation, it may not be sufficient to survive climate change under predicted sea surface temperature scenarios over the next 100 years. However, it may be enough to 'buy time' while greenhouse reduction measures are put in place.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Anthony K.R.N, Hoegh-Guldberg O. Kinetics of photoacclimation in corals. Oecologia. 2003;134:23–31. doi:10.1007/s00442-002-1095-1 - DOI - PubMed
-
- Baker A.C. Reef corals bleach to survive change. Nature. 2001;411:765–766. doi:10.1038/35081151 - DOI - PubMed
-
- Baker A.C. Flexibility and specificity in coral–algal symbiosis: diversity, ecology, and biogeography of Symbiodinium. Annu. Rev. Ecol. Syst. 2003;34:661–689. doi:10.1146/annurev.ecolsys.34.011802.132417 - DOI
-
- Baker A.C, Starger C.J, McClanahan T.R, Glynn P.W. Corals' adaptive response to climate. Nature. 2004;430:741. doi:10.1038/430741a - DOI - PubMed
-
- Berkelmans R, De'ath G, Kininmonth S, Skirving W.J. A comparison of the 1998 and 2002 coral bleaching events on the Great Barrier Reef: spatial correlation, patterns and predictions. Coral Reefs. 2004;23:74–83. doi:10.1007/s00338-003-0353-y - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials