Tollip regulates proinflammatory responses to interleukin-1 and lipopolysaccharide
- PMID: 16428431
- PMCID: PMC1347014
- DOI: 10.1128/MCB.26.3.735-742.2006
Tollip regulates proinflammatory responses to interleukin-1 and lipopolysaccharide
Abstract
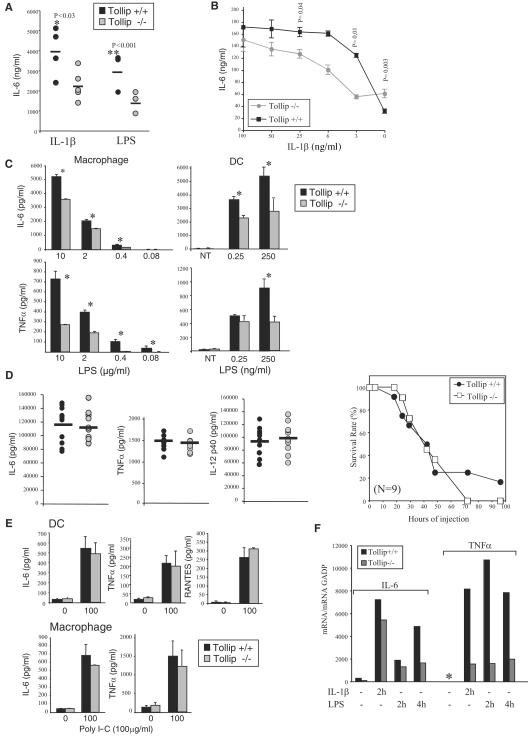
Activation of interleukin-1 (IL-1) receptor (IL-1R), Toll-like receptor 2 (TLR2), and TLR4 triggers NF-kappaB and mitogen-activated protein kinase (MAPK)-dependent signaling, thereby initiating immune responses. Tollip has been implicated as a negative regulator of NF-kappaB signaling triggered by these receptors in in vitro studies. Here, deficient mice were used to determine the physiological contribution of Tollip to immunity. NF-kappaB, as well as MAPK, signaling appeared normal in Tollip-deficient cells stimulated with IL-1beta or the TLR4 ligand lipopolysaccharide (LPS). Similarly, IL-1beta- and TLR-driven activation of dendritic cells and lymphocytes was indistinguishable from wild-type cells. In contrast, the production of the proinflammatory cytokines, IL-6 and tumor necrosis factor alpha was significantly reduced after IL-1beta and LPS treatment at low doses but not at lethal doses of LPS. Tollip therefore controls the magnitude of inflammatory cytokine production in response to IL-1beta and LPS.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Akira, S., and K. Takeda. 2004. Toll-like receptor signalling. Nat. Rev. Immunol. 4:499-511. - PubMed
-
- Bulut, Y., E. Faure, L. Thomas, O. Equils, and M. Arditi. 2001. Cooperation of Toll-like receptor 2 and 6 for cellular activation by soluble tuberculosis factor and Borrelia burgdorferi outer surface protein A lipoprotein: role of Toll-interacting protein and IL-1 receptor signaling molecules in Toll-like receptor 2 signaling. J. Immunol. 167:987-994. - PubMed
-
- Burns, K., J. Clatworthy, L. Martin, F. Martinon, C. Plumpton, B. Maschera, A. Lewis, K. Ray, J. Tschopp, and F. Volpe. 2000. Tollip, a new component of the IL-1RI pathway, links IRAK to the IL-1 receptor. Nat. Cell Biol. 2:346-351. - PubMed
-
- Didierlaurent, A., I. Ferrero, L. A. Otten, B. Dubois, M. Reinhardt, H. Carlsen, R. Blomhoff, S. Akira, J. P. Kraehenbuhl, and J. C. Sirard. 2004. Flagellin promotes myeloid differentiation factor 88-dependent development of Th2-type response. J. Immunol. 172:6922-6930. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases