The interaction site of Flap Endonuclease-1 with WRN helicase suggests a coordination of WRN and PCNA
- PMID: 16326861
- PMCID: PMC1301591
- DOI: 10.1093/nar/gki1002
The interaction site of Flap Endonuclease-1 with WRN helicase suggests a coordination of WRN and PCNA
Abstract
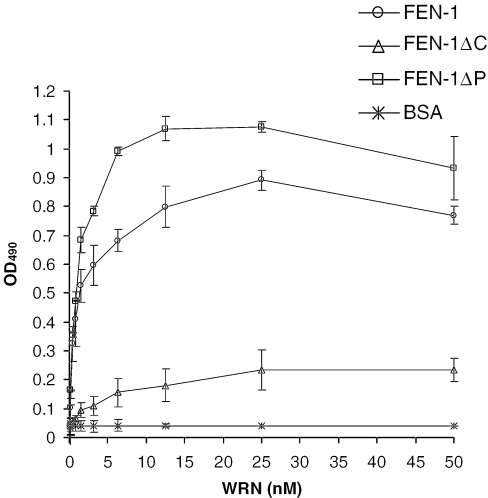
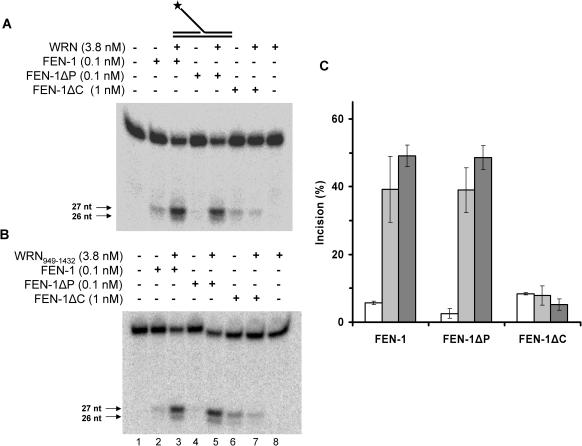
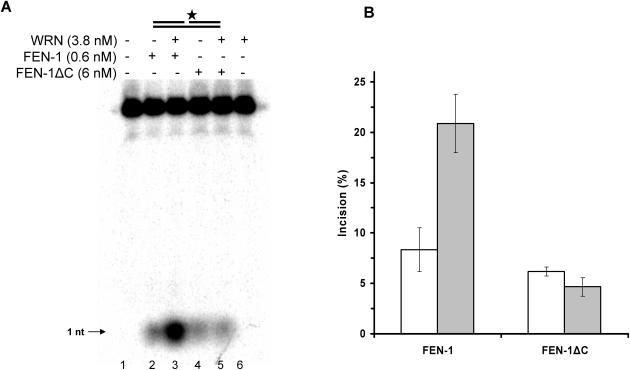
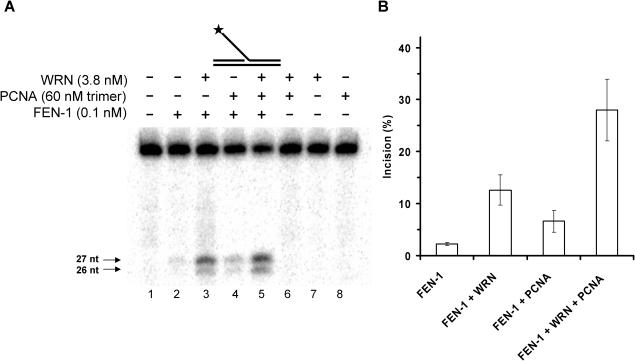
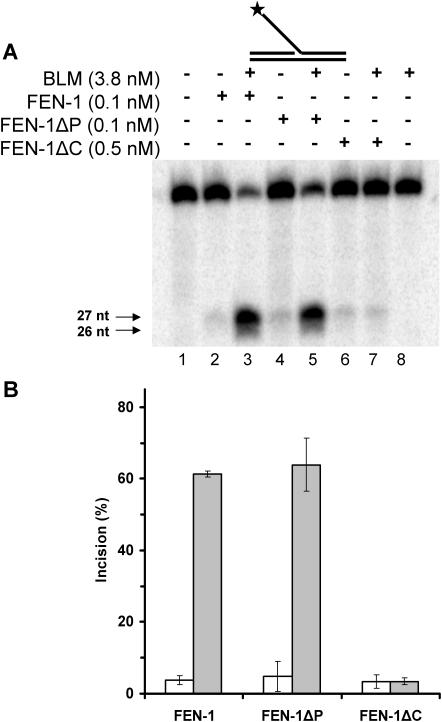
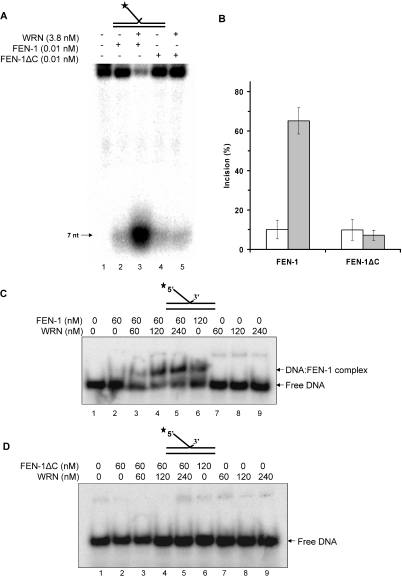
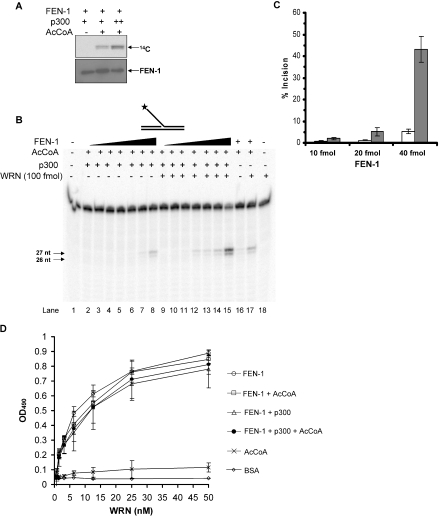
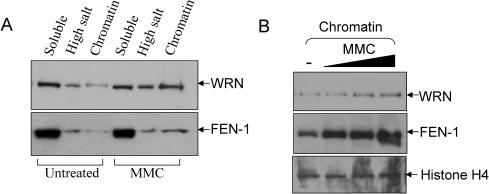
Werner and Bloom syndromes are genetic RecQ helicase disorders characterized by genomic instability. Biochemical and genetic data indicate that an important protein interaction of WRN and Bloom syndrome (BLM) helicases is with the structure-specific nuclease Flap Endonuclease 1 (FEN-1), an enzyme that is implicated in the processing of DNA intermediates that arise during cellular DNA replication, repair and recombination. To acquire a better understanding of the interaction of WRN and BLM with FEN-1, we have mapped the FEN-1 binding site on the two RecQ helicases. Both WRN and BLM bind to the extreme C-terminal 18 amino acid tail of FEN-1 that is adjacent to the PCNA binding site of FEN-1. The importance of the WRN/BLM physical interaction with the FEN-1 C-terminal tail was confirmed by functional interaction studies with catalytically active purified recombinant FEN-1 deletion mutant proteins that lack either the WRN/BLM binding site or the PCNA interaction site. The distinct binding sites of WRN and PCNA and their combined effect on FEN-1 nuclease activity suggest that they may coordinately act with FEN-1. WRN was shown to facilitate FEN-1 binding to its preferred double-flap substrate through its protein interaction with the FEN-1 C-terminal binding site. WRN retained its ability to physically bind and stimulate acetylated FEN-1 cleavage activity to the same extent as unacetylated FEN-1. These studies provide new insights to the interaction of WRN and BLM helicases with FEN-1, and how these interactions might be regulated with the PCNA-FEN-1 interaction during DNA replication and repair.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Martin G.M. Genetic syndromes in man with potential relevance to the pathobiology of aging. Birth Defects Orig. Artic. Ser. 1978;14:5–15. - PubMed
-
- Hanaoka F., Yamada M., Takeuchi F., Goto M., Miyamoto T., Hori T. Autoradiographic studies of DNA replication in Werner's syndrome cells. Adv. Exp. Med. Biol. 1985;190:439–457. - PubMed
-
- Martin G.M., Sprague C.A., Epstein C.J. Replicative life-span of cultivated human cells. Effects of donor's age, tissue, and genotype. Lab. Invest. 1970;23:86–92. - PubMed
-
- Poot M., Hoehn H., Runger T.M., Martin G.M. Impaired S-phase transit of Werner syndrome cells expressed in lymphoblastoid cells. Exp. Cell. Res. 1992;202:267–273. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous