Inhibition of alpha/beta interferon signaling by the NS4B protein of flaviviruses
- PMID: 15956546
- PMCID: PMC1143737
- DOI: 10.1128/JVI.79.13.8004-8013.2005
Inhibition of alpha/beta interferon signaling by the NS4B protein of flaviviruses
Abstract
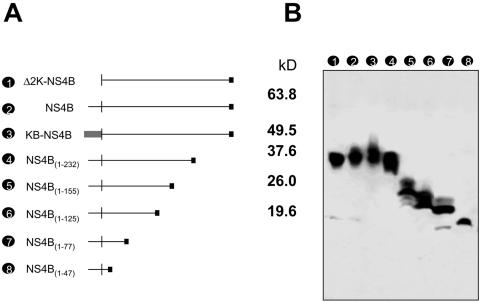
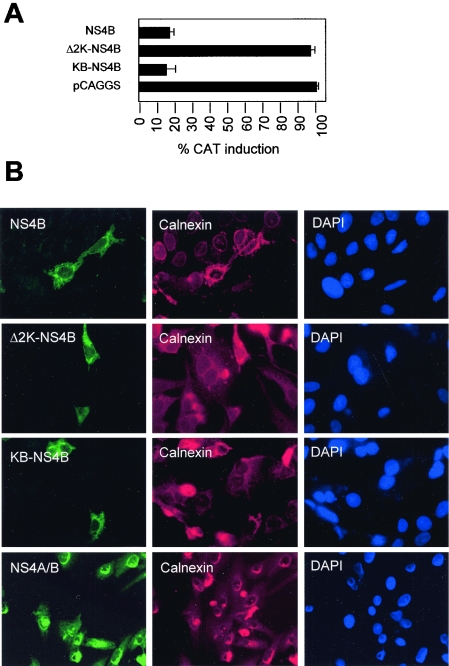
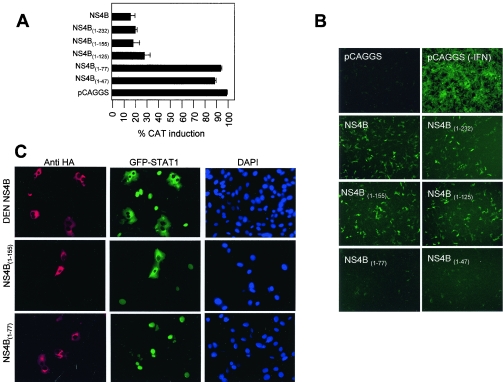
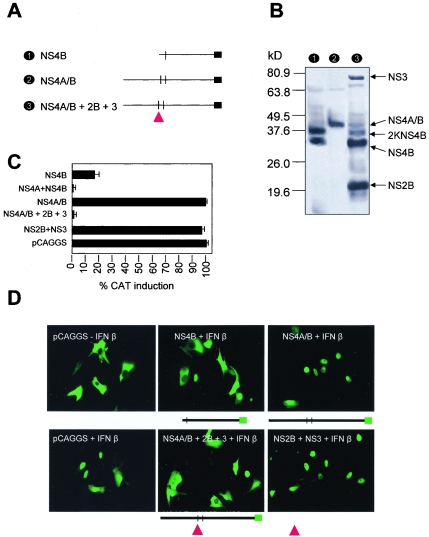
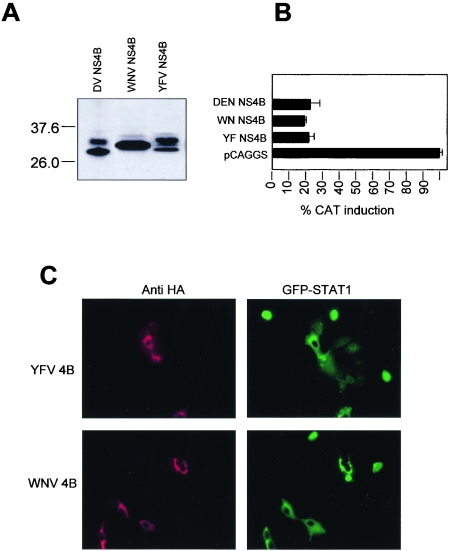
Flaviviruses are insect-borne, positive-strand RNA viruses that have been disseminated worldwide. Their genome is translated into a polyprotein, which is subsequently cleaved by a combination of viral and host proteases to produce three structural proteins and seven nonstructural proteins. The nonstructural protein NS4B of dengue 2 virus partially blocks activation of STAT1 and interferon-stimulated response element (ISRE) promoters in cells stimulated with interferon (IFN). We have found that this function of NS4B is conserved in West Nile and yellow fever viruses. Deletion analysis shows that that the first 125 amino acids of dengue virus NS4B are sufficient for inhibition of alpha/beta IFN (IFN-alpha/beta) signaling. The cleavable signal peptide at the N terminus of NS4B, a peptide with a molecular weight of 2,000, is required for IFN antagonism but can be replaced by an unrelated signal peptide. Coexpression of dengue virus NS4A and NS4B together results in enhanced inhibition of ISRE promoter activation in response to IFN-alpha/beta. In contrast, expression of the precursor NS4A/B fusion protein does not cause an inhibition of IFN signaling unless this product is cleaved by the viral peptidase NS2B/NS3, indicating that proper viral polyprotein processing is required for anti-interferon function.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Chu, P. W., and E. G. Westaway. 1992. Molecular and ultrastructural analysis of heavy membrane fractions associated with the replication of Kunjin virus RNA. Arch. Virol. 125:177-191. - PubMed
-
- Deblandre, G. A., O. P. Marinx, S. S. Evans, S. Majjaj, O. Leo, D. Caput, G. A. Huez, and M. G. Wathelet. 1995. Expression cloning of an interferon-inducible 17-kDa membrane protein implicated in the control of cell growth. J. Biol. Chem. 270:23860-23866. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous