Toll-like receptor 4 contributes to efficient control of infection with the protozoan parasite Leishmania major
- PMID: 15039311
- PMCID: PMC375159
- DOI: 10.1128/IAI.72.4.1920-1928.2004
Toll-like receptor 4 contributes to efficient control of infection with the protozoan parasite Leishmania major
Abstract
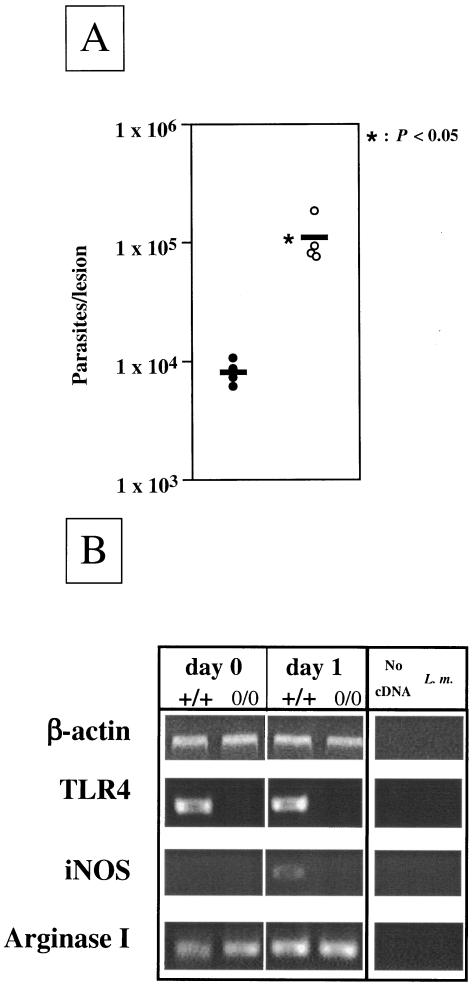
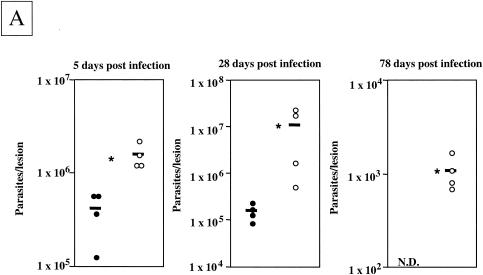
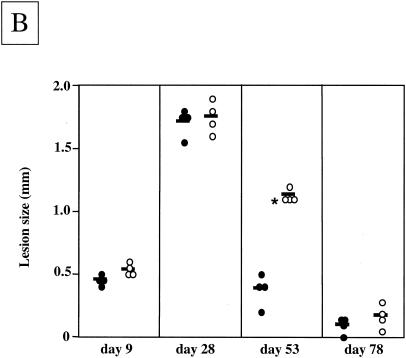
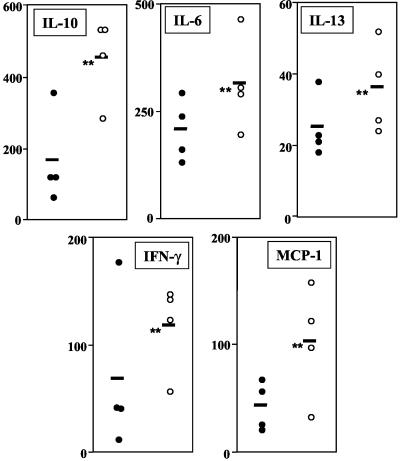
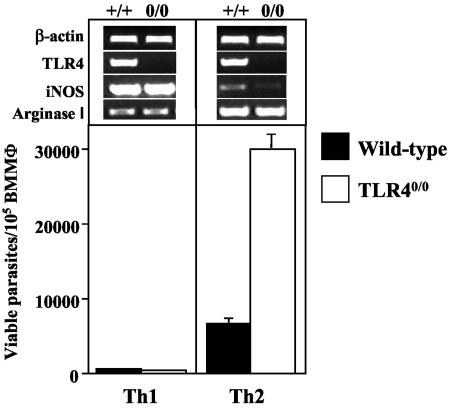
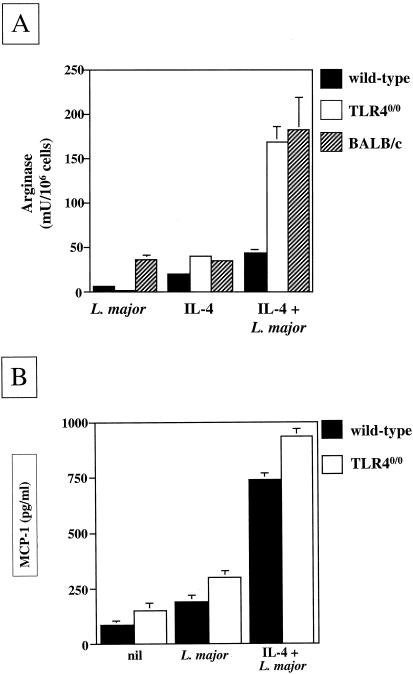
The essential role of Toll-like receptors (TLR) in innate immune responses to bacterial pathogens is increasingly recognized, but very little is known about the role of TLRs in host defense against infections with eukaryotic pathogens. For the present study, we investigated whether TLRs contribute to the innate and acquired immune response to infection with the intracellular protozoan parasite Leishmania major. Our results show that TLR4 contributes to the control of parasite growth in both phases of the immune response. We also addressed the mechanism that results in killing or growth of the intracellular parasites. Control of parasite replication correlates with the early induction of inducible nitric oxide synthase in TLR4-competent mice, whereas increased parasite survival in host cells from TLR4-deficient mice correlates with a higher activity of arginase, an enzyme known to promote parasite growth. This is the first study showing that TLR4 contributes to the effective control of Leishmania infection in vivo.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Aliprantis, A. O., R.-B. Yang, M. R. Mark, S. Suggett, B. Devaux, J. D. Radolf, G. R. Klimpel, P. Godowski, and A. Zychlinsky. 1999. Cell activation and apoptosis by bacterial lipoproteins through Toll-like receptor-2. Science 285:736-739. - PubMed
-
- Belkaid, Y., K. F. Hoffmann, S. Mendez, S. Kamhawi, M. C. Udey, T. A. Wynn, and D. L. Sacks. 2001. The role of interleukin (IL)-10 in the persistence of Leishmania major in the skin after healing and the therapeutic potential of anti-IL-10 receptor antibody for sterile cure. J. Exp. Med. 194:1497-1506. - PMC - PubMed
-
- Bogdan, C. 2000. The function of type I interferons in antimicrobial immunity. Curr. Opin. Immunol. 12:419-424. - PubMed
-
- Bogdan, C. 2001. Nitric oxide and the immune response. Nat. Immunol. 2:907-916. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases