Brucella evades macrophage killing via VirB-dependent sustained interactions with the endoplasmic reticulum
- PMID: 12925673
- PMCID: PMC2194179
- DOI: 10.1084/jem.20030088
Brucella evades macrophage killing via VirB-dependent sustained interactions with the endoplasmic reticulum
Abstract
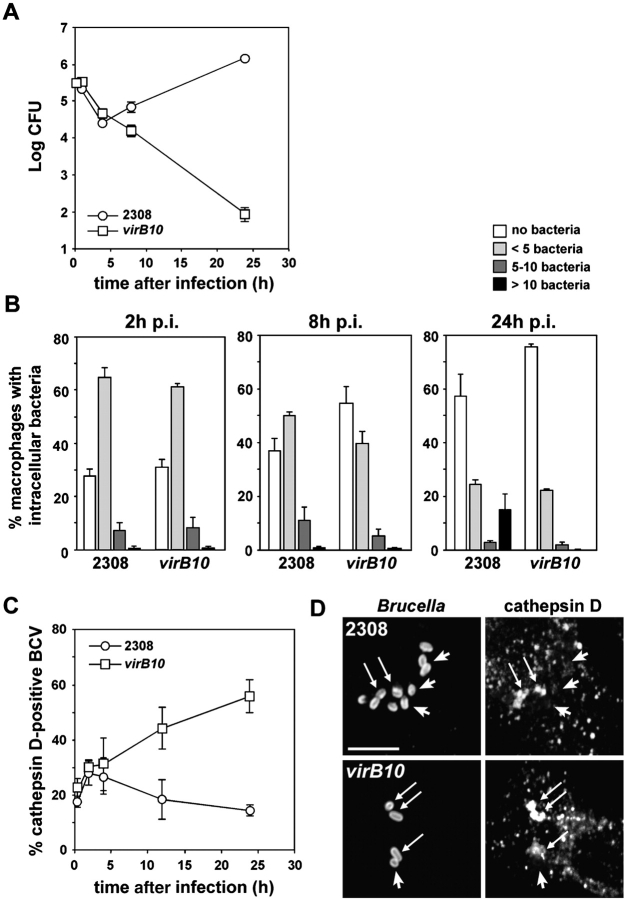
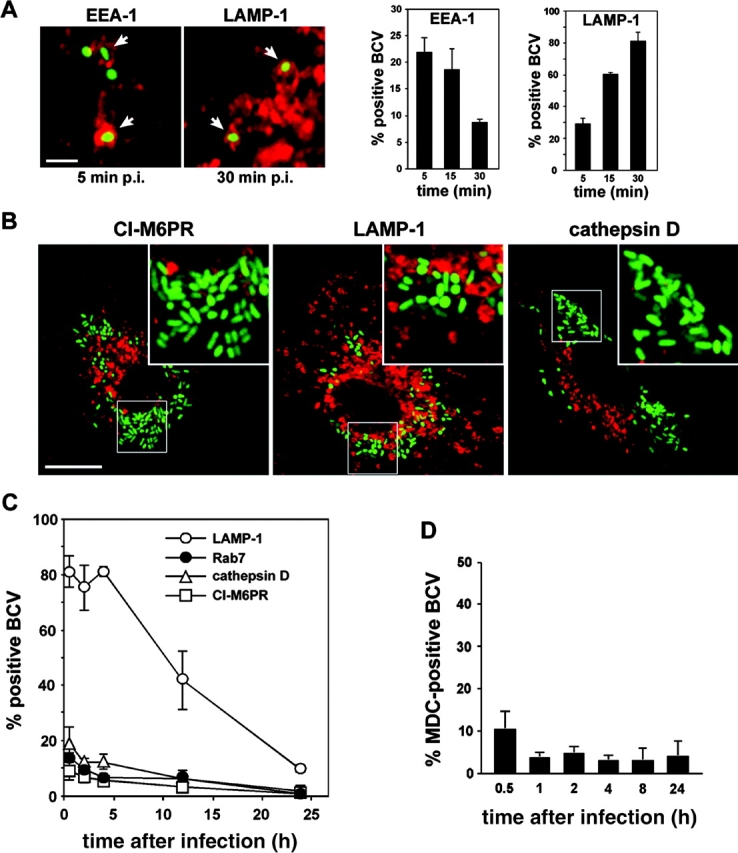
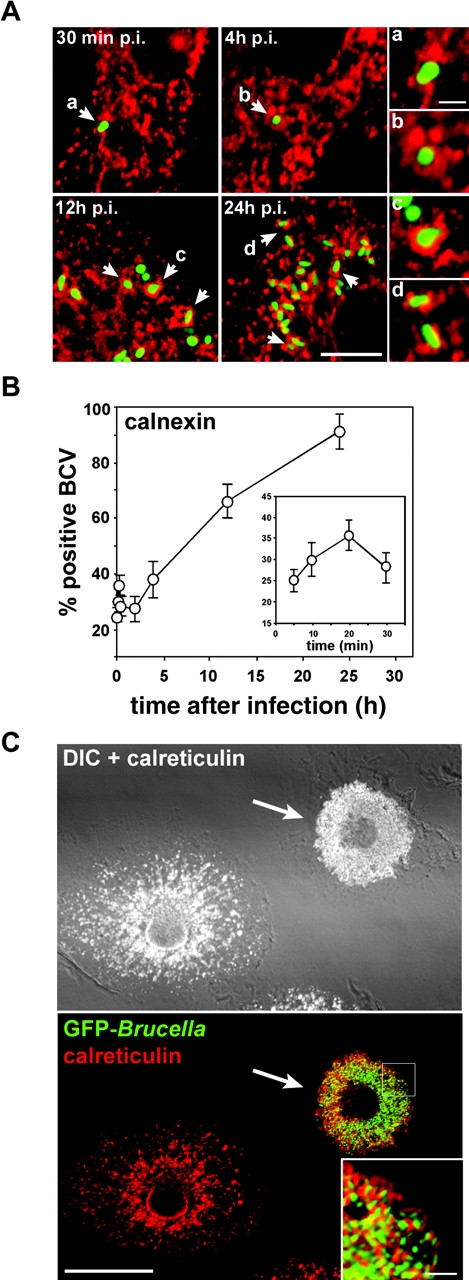
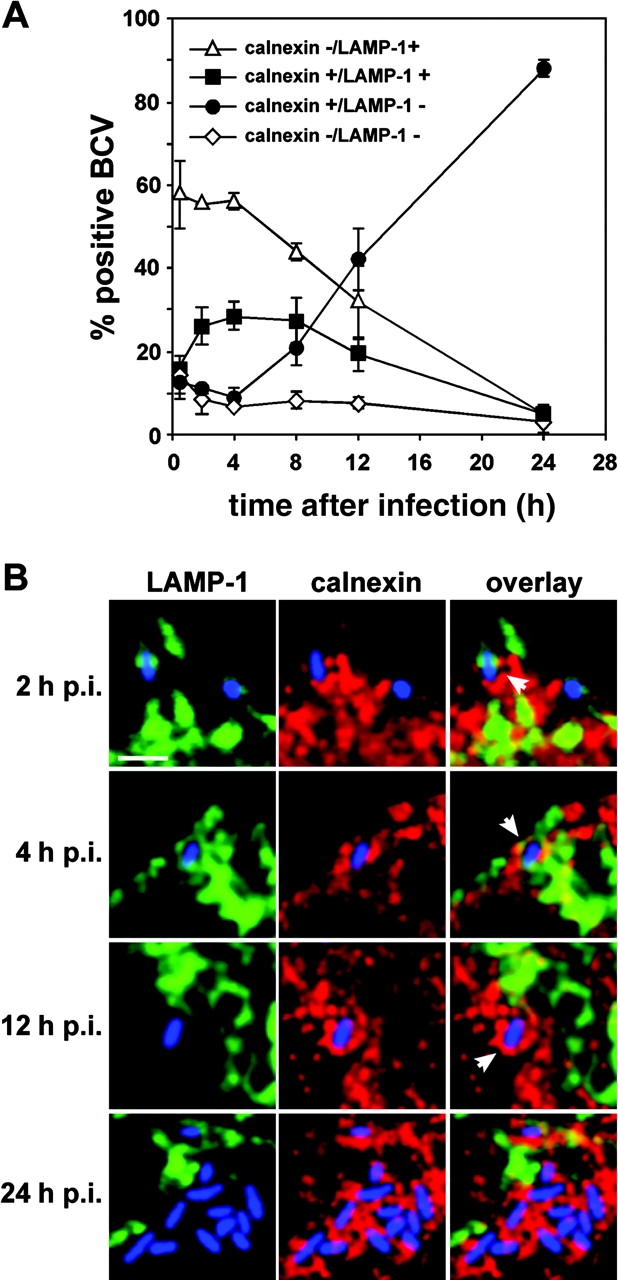
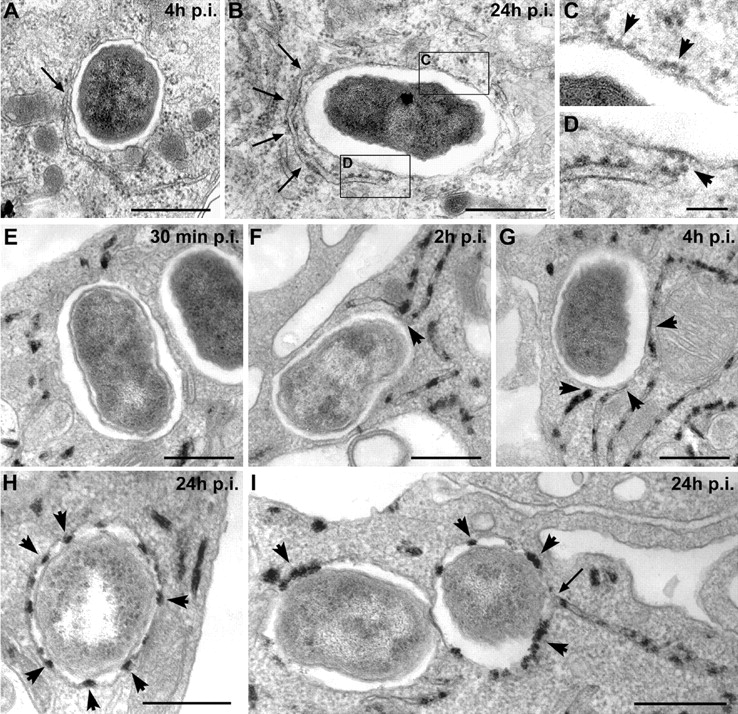
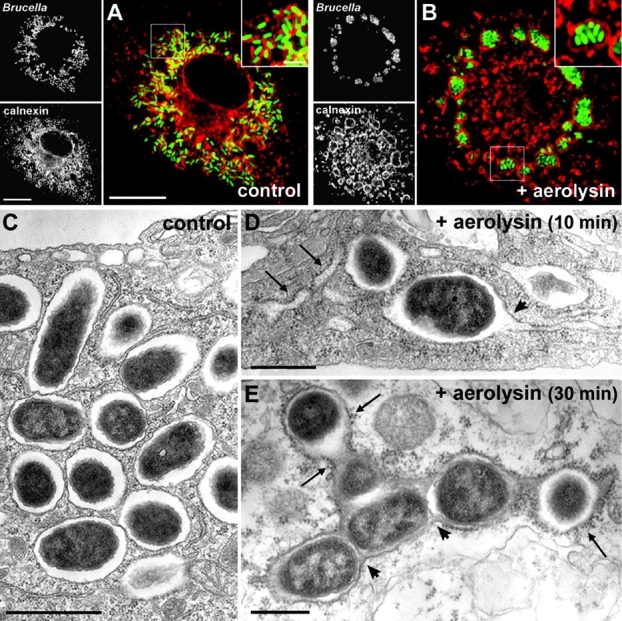
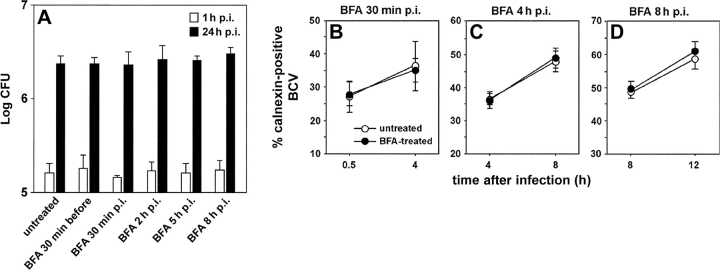
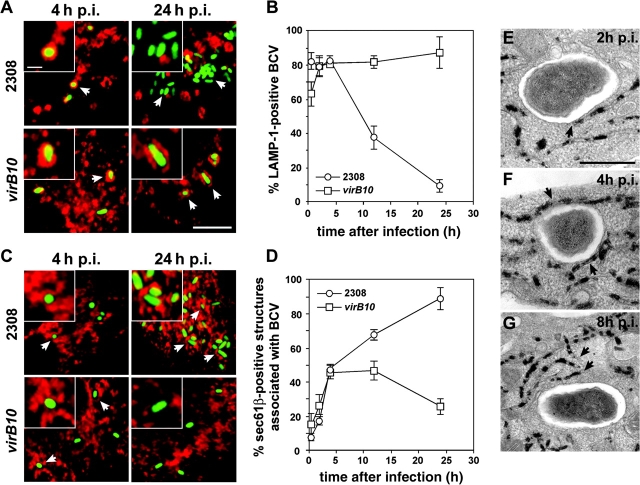
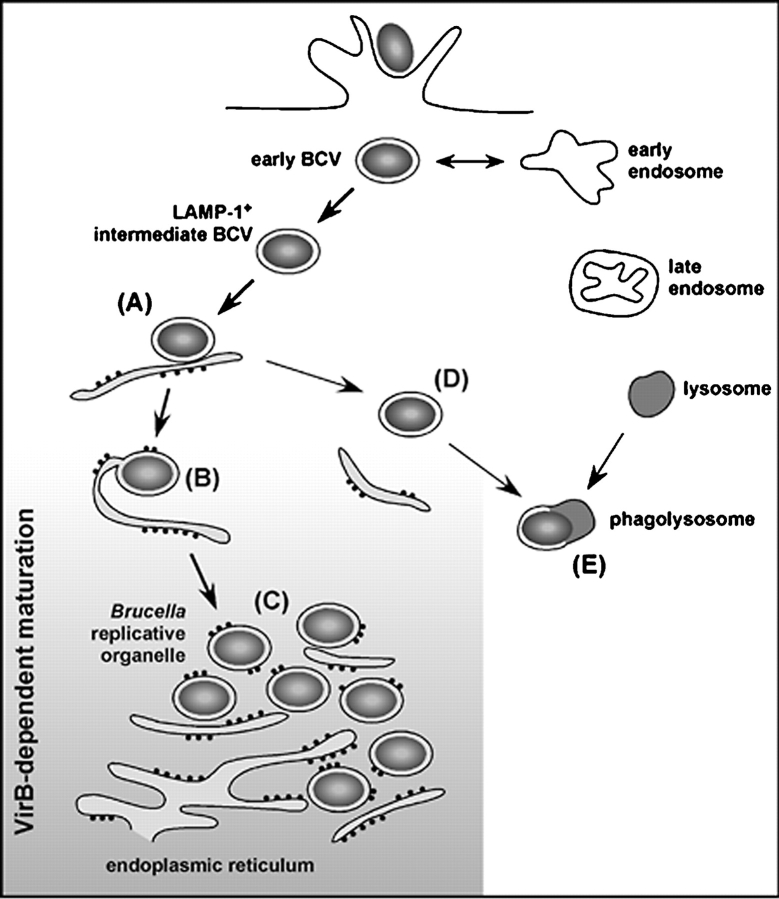
The intracellular pathogen Brucella is the causative agent of brucellosis, a worldwide zoonosis that affects mammals, including humans. Essential to Brucella virulence is its ability to survive and replicate inside host macrophages, yet the underlying mechanisms and the nature of the replicative compartment remain unclear. Here we show in a model of Brucella abortus infection of murine bone marrow-derived macrophages that a fraction of the bacteria that survive an initial macrophage killing proceed to replicate in a compartment segregated from the endocytic pathway. The maturation of the Brucella-containing vacuole involves sustained interactions and fusion with the endoplasmic reticulum (ER), which creates a replicative compartment with ER-like properties. The acquisition of ER membranes by replicating Brucella is independent of ER-Golgi COPI-dependent vesicular transport. A mutant of the VirB type IV secretion system, which is necessary for intracellular survival, was unable to sustain interactions and fuse with the ER, and was killed via eventual fusion with lysosomes. Thus, we demonstrate that live intracellular Brucella evade macrophage killing through VirB-dependent sustained interactions with the ER. Moreover, we assign an intracellular function to the VirB system, as being required for late maturation events necessary for the biogenesis of an ER-derived replicative organelle.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Knodler, L.A., J. Celli, and B.B. Finlay. 2001. Pathogenic trickery: deception of host cell processes. Nat. Rev. Mol. Cell Biol. 2:578–588. - PubMed
-
- Moreno, E., and I. Moriyon. 2001. The genus Brucella. The Prokaryotes. M. Dorkin, S. Falkow, E. Rosenberg, K.-H. Schleifer, and E. Stackebrandt, editors. Springer-Verlag, New York.
-
- Gorvel, J.P., and E. Moreno. 2002. Brucella intracellular life: from invasion to intracellular replication. Vet. Microbiol. 90:281–297. - PubMed