Multicellularity and increasing Reynolds number impact on the evolutionary shift in flash-induced ciliary response in Volvocales
- PMID: 39277710
- PMCID: PMC11401373
- DOI: 10.1186/s12862-024-02307-1
Multicellularity and increasing Reynolds number impact on the evolutionary shift in flash-induced ciliary response in Volvocales
Abstract
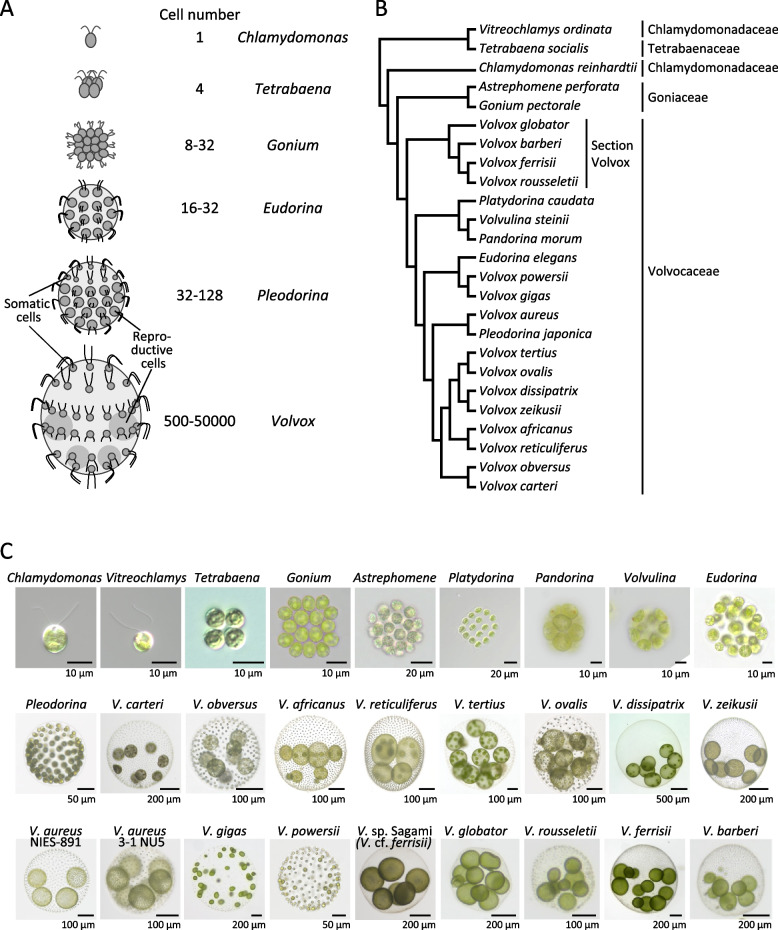
Background: Volvocales in green algae have evolved by multicellularity of Chlamydomonas-like unicellular ancestor. Those with various cell numbers exist, such as unicellular Chlamydomonas, four-celled Tetrabaena, and Volvox species with different cell numbers (~1,000, ~5,000, and ~10,000). Each cell of these organisms shares two cilia and an eyespot, which are used for swimming and photosensing. They are all freshwater microalgae but inhabit different fluid environments: unicellular species live in low Reynolds-number (Re) environments where viscous forces dominate, whereas multicellular species live in relatively higher Re where inertial forces become non-negligible. Despite significant changes in the physical environment, during the evolution of multicellularity, they maintained photobehaviors (i.e., photoshock and phototactic responses), which allows them to survive under changing light conditions.
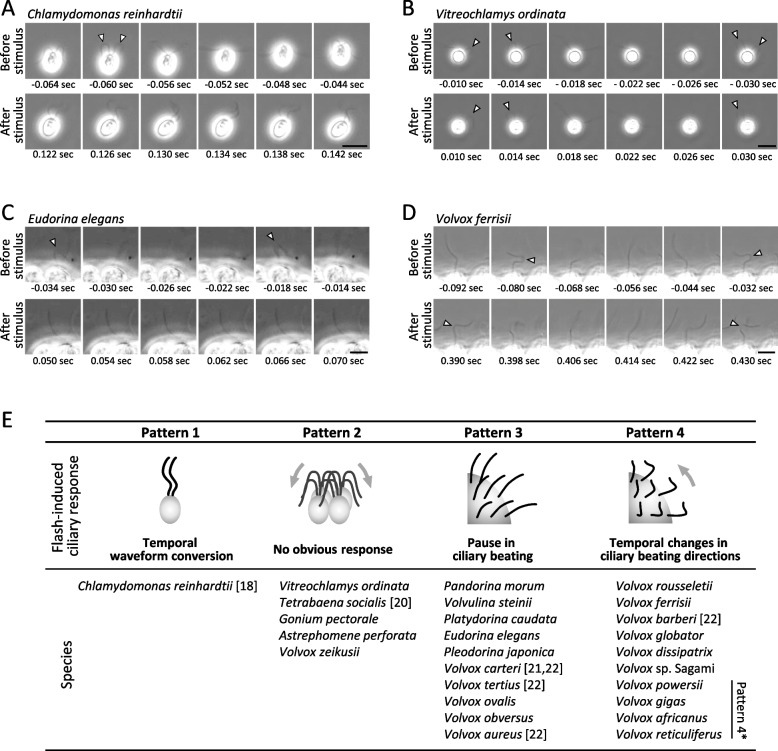
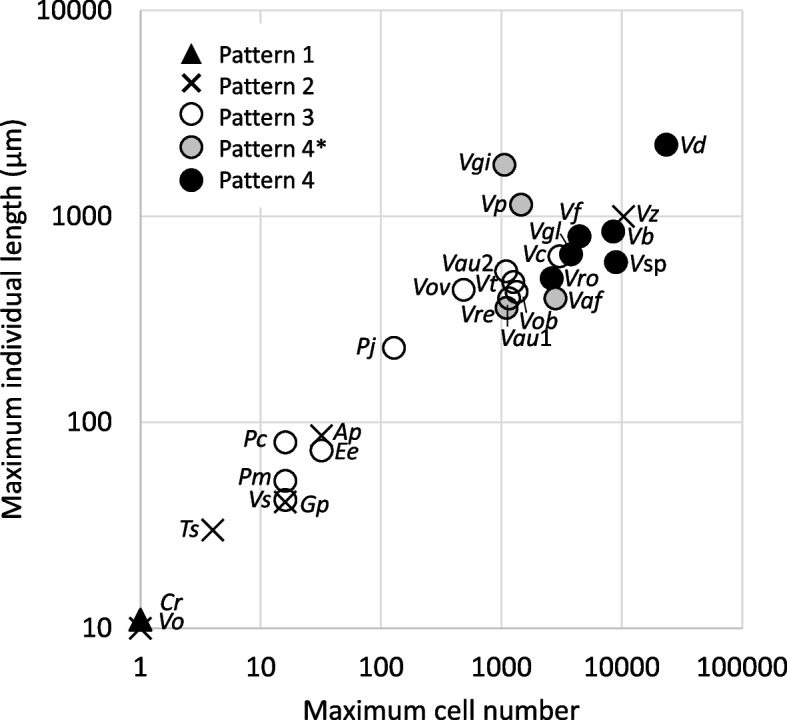
Results: In this study, we utilized high-speed imaging to observe flash-induced changes in the ciliary beating manner of 27 Volvocales strains. We classified flash-induced ciliary responses in Volvocales into four patterns: "1: temporal waveform conversion", "2: no obvious response", "3: pause in ciliary beating", and "4: temporal changes in ciliary beating directions". We found that which species exhibit which pattern depends on Re, which is associated with the individual size of each species rather than phylogenetic relationships.
Conclusions: These results suggest that only organisms that acquired different patterns of ciliary responses survived the evolutionary transition to multicellularity with a greater number of cells while maintaining photobehaviors. This study highlights the significance of the Re as a selection pressure in evolution and offers insights for designing propulsion systems in biomimetic micromachines.
Keywords: Cilia; Flagella; Multicellularity; Reynolds number; Volvocales.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Kirk DL. Volvox: a search for the molecular and genetic origins of multicellularity and cellular differentiation. Cambridge: Cambridge University Press; 1998.
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials