Widespread Sterol Methyltransferase Participates in the Biosynthesis of Both C4α- and C4β-Methyl Sterols
- PMID: 35561259
- PMCID: PMC9136925
- DOI: 10.1021/jacs.2c01401
Widespread Sterol Methyltransferase Participates in the Biosynthesis of Both C4α- and C4β-Methyl Sterols
Abstract
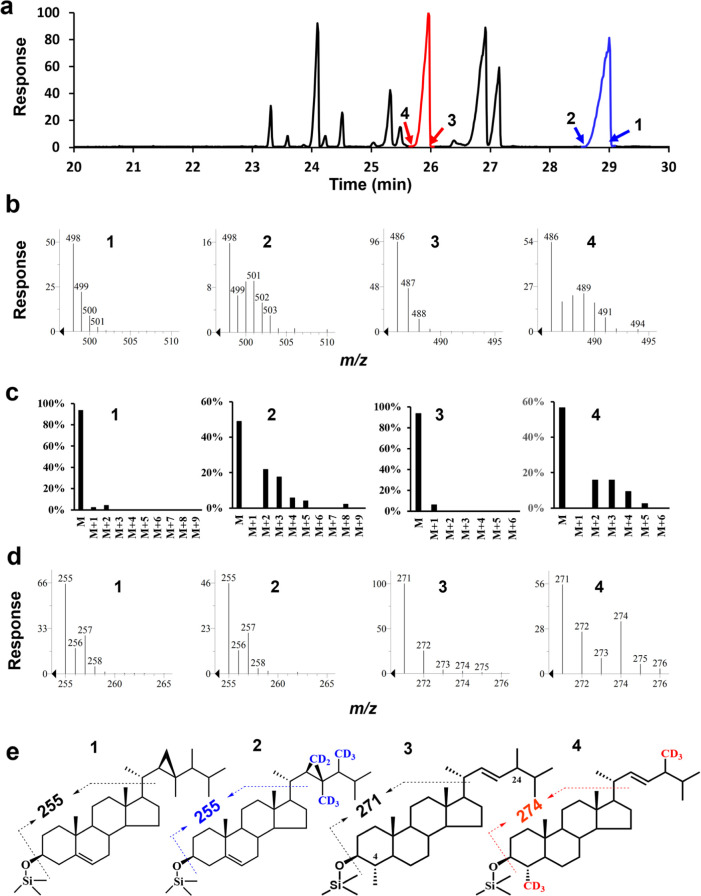
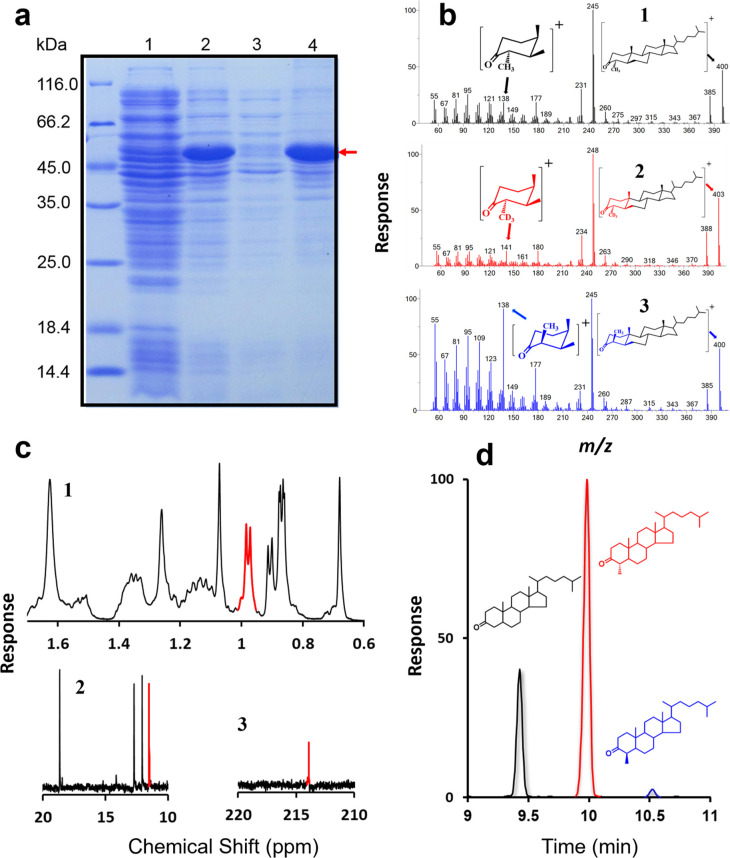
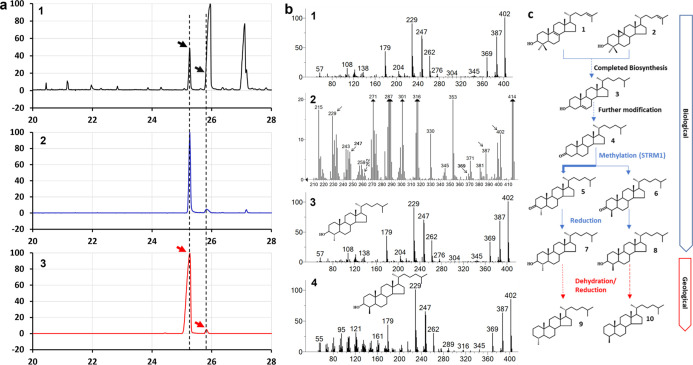
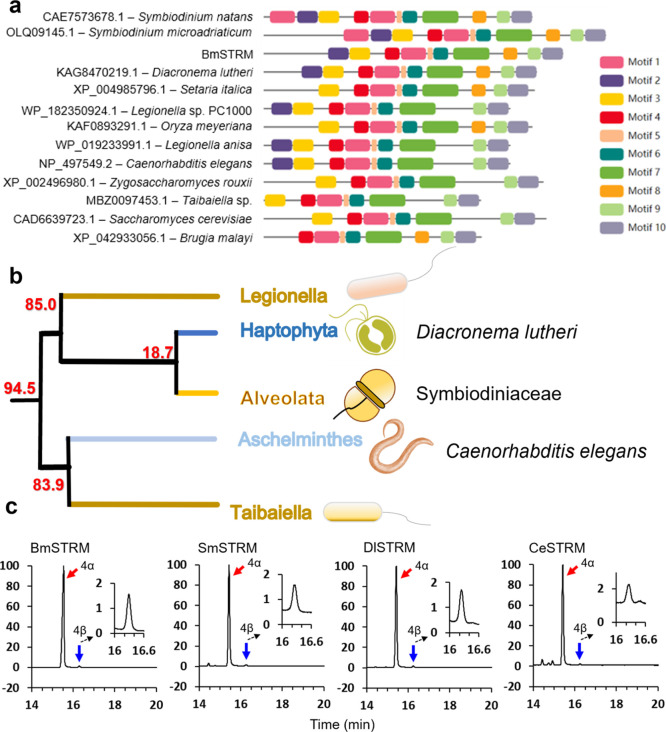
The 4-methyl steranes serve as molecular fossils and are used for studying both eukaryotic evolution and geological history. The occurrence of 4α-methyl steranes in sediments has long been considered evidence of products of partial demethylation mediated by sterol methyl oxidases (SMOs), while 4β-methyl steranes are attributed entirely to diagenetic generation from 4α-methyl steroids since possible biological sources of their precursor 4β-methyl sterols are unknown. Here, we report a previously unknown C4-methyl sterol biosynthetic pathway involving a sterol methyltransferase rather than the SMOs. We show that both C4α- and C4β-methyl sterols are end products of the sterol biosynthetic pathway in an endosymbiont of reef corals, Breviolum minutum, while this mechanism exists not only in dinoflagellates but also in eukaryotes from alveolates, haptophytes, and aschelminthes. Our discovery provides a previously untapped route for the generation of C4-methyl steranes and overturns the paradigm that all 4β-methyl steranes are diagenetically generated from the 4α isomers. This may facilitate the interpretation of molecular fossils and understanding of the evolution of eukaryotic life in general.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Goad L. J. The biosynthesis of plant sterols. Lipids Lipid Polym. Higher Plants, [Pap. Symp.] 1977, 146–168. 10.1007/978-3-642-66632-2_8. - DOI
-
- Robinson N.; Eglinton G.; Brassell S. C.; Cranwell P. A. Dinoflagellate origin for sedimentary 4α-methylsteroids and 5α(H)-stanols. Nature 1984, 308, 439–442. 10.1038/308439a0. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous