The four-celled Volvocales green alga Tetrabaena socialis exhibits weak photobehavior and high-photoprotection ability
- PMID: 34699573
- PMCID: PMC8547699
- DOI: 10.1371/journal.pone.0259138
The four-celled Volvocales green alga Tetrabaena socialis exhibits weak photobehavior and high-photoprotection ability
Abstract
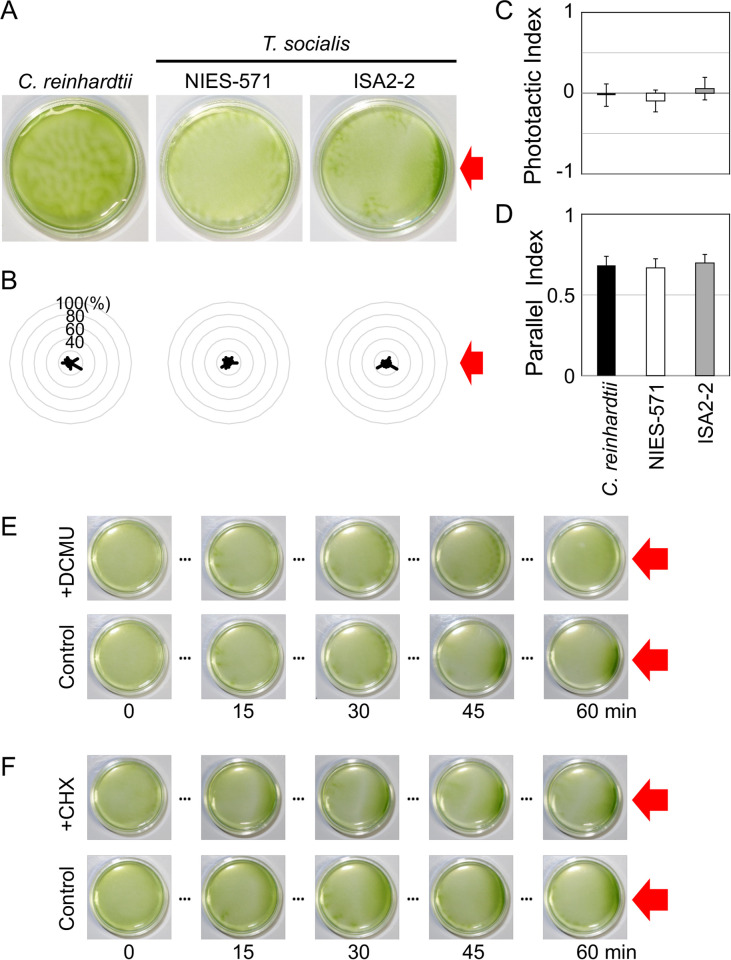
Photo-induced behavioral responses (photobehaviors) are crucial to the survival of motile phototrophic organisms in changing light conditions. Volvocine green algae are excellent model organisms for studying the regulatory mechanisms of photobehavior. We recently reported that unicellular Chlamydomonas reinhardtii and multicellular Volvox rousseletii exhibit similar photobehaviors, such as phototactic and photoshock responses, via different ciliary regulations. To clarify how the regulatory systems have changed during the evolution of multicellularity, we investigated the photobehaviors of four-celled Tetrabaena socialis. Surprisingly, unlike C. reinhardtii and V. rousseletii, T. socialis did not exhibit immediate photobehaviors after light illumination. Electrophysiological analysis revealed that the T. socialis eyespot does not function as a photoreceptor. Instead, T. socialis exhibited slow accumulation toward the light source in a photosynthesis-dependent manner. Our assessment of photosynthetic activities showed that T. socialis chloroplasts possess higher photoprotection abilities against strong light than C. reinhardtii. These data suggest that C. reinhardtii and T. socialis employ different strategies to avoid high-light stress (moving away rapidly and gaining photoprotection, respectively) despite their close phylogenetic relationship.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Demmig-Adams B, Adams WW, 3rd. Photoprotection and other responses of plants to high light stress. Annu Rev Plant Physiol Plant Mol Bioi 1992. p. 599–626.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials