Guanine, a high-capacity and rapid-turnover nitrogen reserve in microalgal cells
- PMID: 33293415
- PMCID: PMC7768779
- DOI: 10.1073/pnas.2005460117
Guanine, a high-capacity and rapid-turnover nitrogen reserve in microalgal cells
Abstract
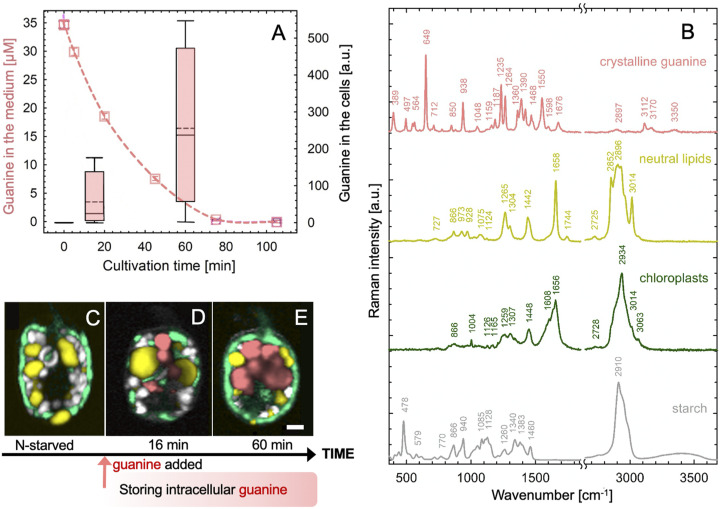
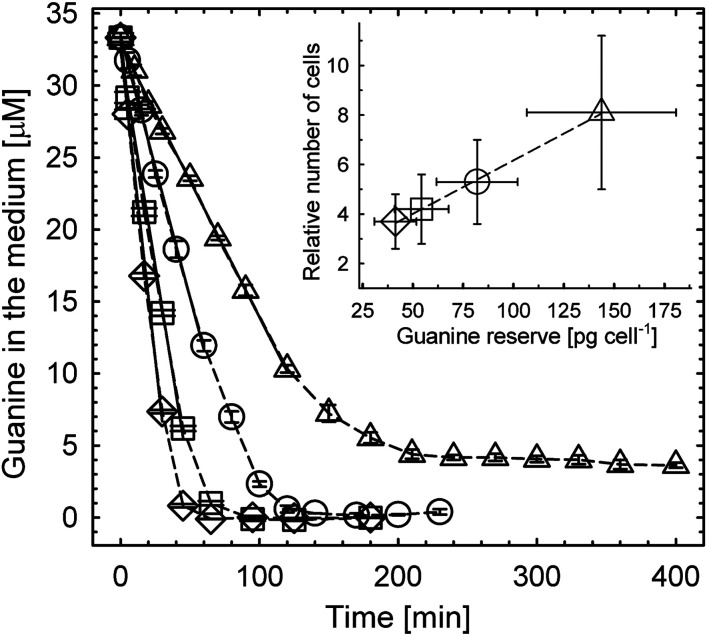
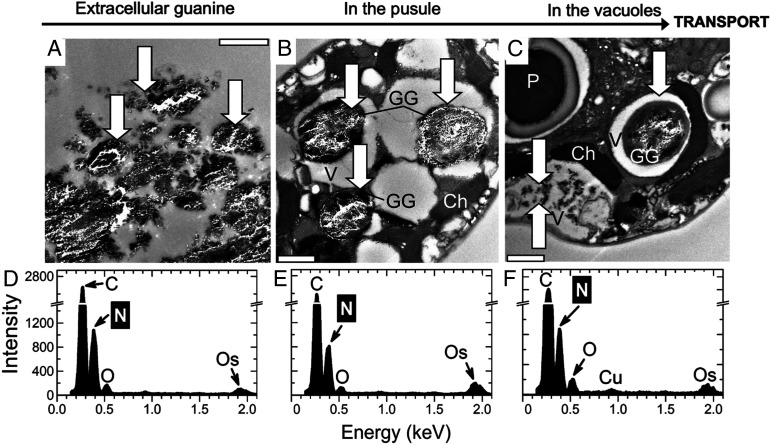
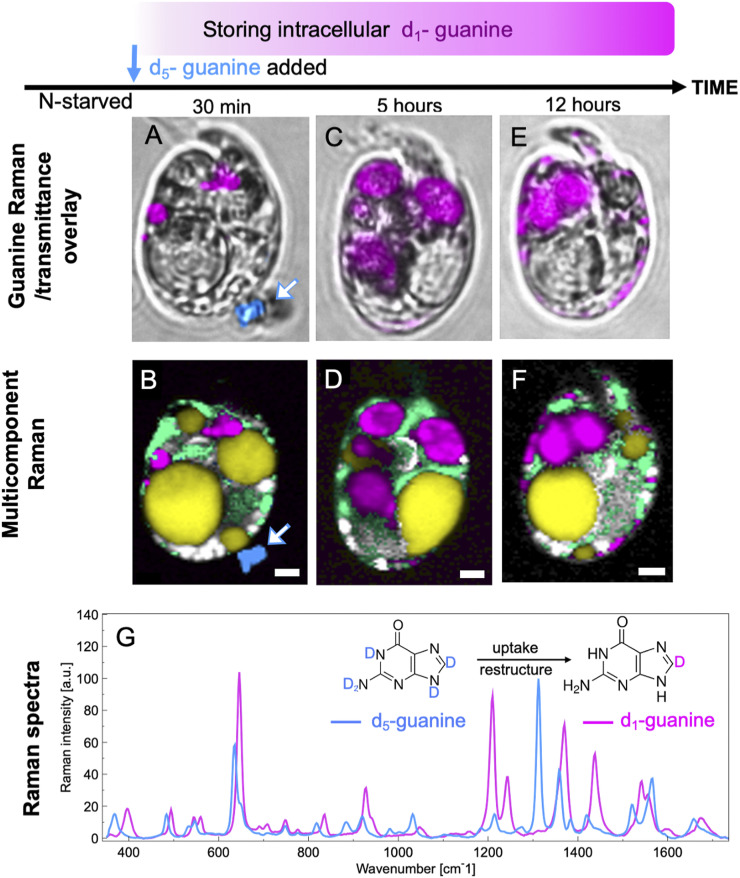
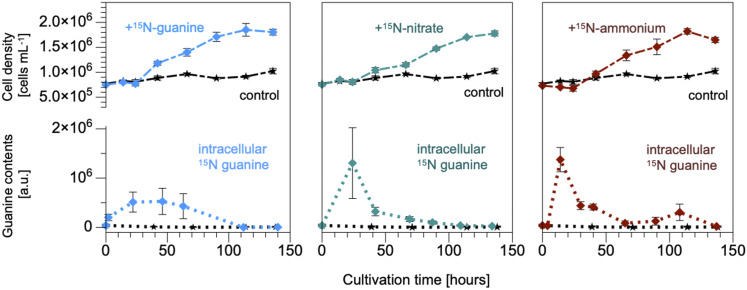
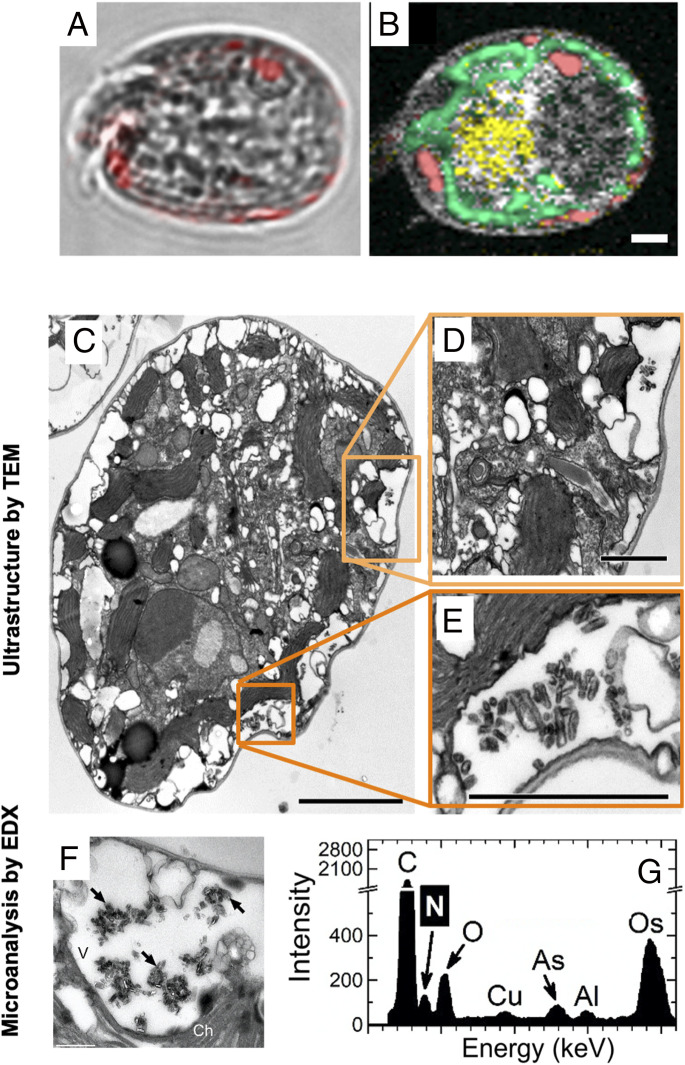
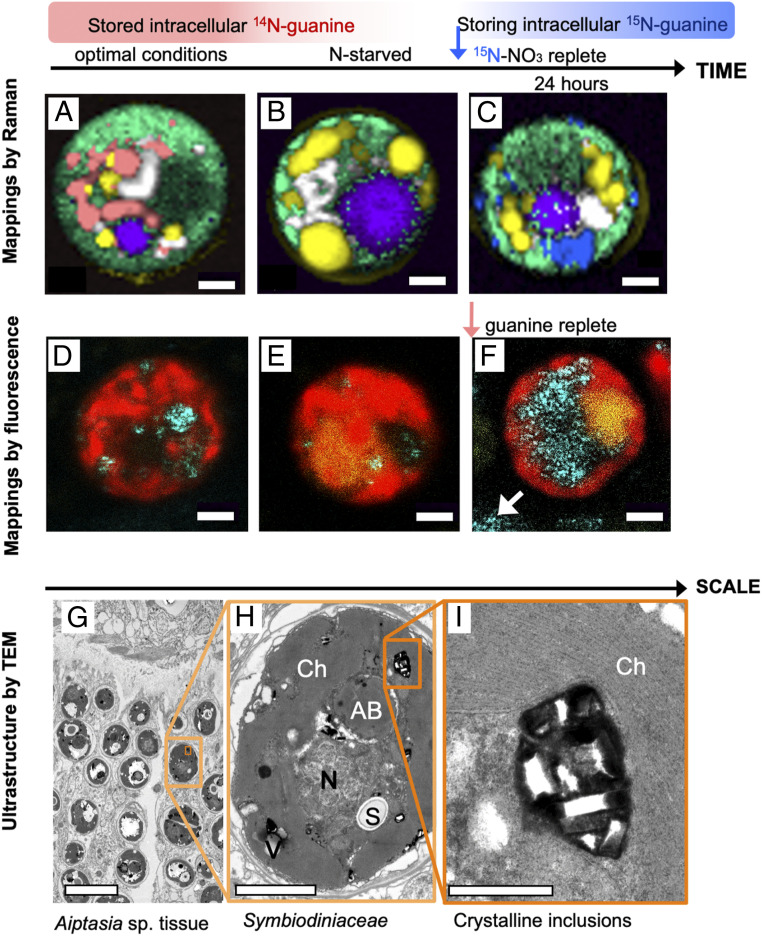
Nitrogen (N) is an essential macronutrient for microalgae, influencing their productivity, composition, and growth dynamics. Despite the dramatic consequences of N starvation, many free-living and endosymbiotic microalgae thrive in N-poor and N-fluctuating environments, giving rise to questions about the existence and nature of their long-term N reserves. Our understanding of these processes requires a unequivocal identification of the N reserves in microalgal cells as well as their turnover kinetics and subcellular localization. Herein, we identified crystalline guanine as the enigmatic large-capacity and rapid-turnover N reserve of microalgae. The identification was unambiguously supported by confocal Raman, fluorescence, and analytical transmission electron microscopies as well as stable isotope labeling. We discovered that the storing capacity for crystalline guanine by the marine dinoflagellate Amphidiniumcarterae was sufficient to support N requirements for several new generations. We determined that N reserves were rapidly accumulated from guanine available in the environment as well as biosynthesized from various N-containing nutrients. Storage of exogenic N in the form of crystalline guanine was found broadly distributed across taxonomically distant groups of microalgae from diverse habitats, from freshwater and marine free-living forms to endosymbiotic microalgae of reef-building corals (Acropora millepora, Euphyllia paraancora). We propose that crystalline guanine is the elusive N depot that mitigates the negative consequences of episodic N shortage. Guanine (C5H5N5O) may act similarly to cyanophycin (C10H19N5O5) granules in cyanobacteria. Considering the phytoplankton nitrogen pool size and dynamics, guanine is proposed to be an important storage form participating in the global N cycle.
Keywords: coral; guanine; nitrogen cycle; nutrient storage; phytoplankton.
Conflict of interest statement
The authors declare no competing interest.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Antia N. J., Harrison P. J., Oliveira L., The role of dissolved organic nitrogen in phytoplankton nutrition, cell biology and ecology. Phycologia 30, 1–89 (1991).
-
- Falkowski P. G., Evolution of the nitrogen cycle and its influence on the biological sequestration of CO2 in the ocean. Nature 387, 272–275 (1997).
-
- Falkowski P. G., Barber R. T., Smetacek V., Biogeochemical controls and feedbacks on ocean primary production. Science 281, 200–207 (1998). - PubMed
-
- Anderson D. M., Glibert P. M., Burkholder J. M., Harmful algal blooms and eutrophication: Nutrient sources, composition, and consequences. Estuaries 25, 704–726 (2002).
-
- Rädecker N., Pogoreutz C., Voolstra C. R., Wiedenmann J., Wild C., Nitrogen cycling in corals: The key to understanding holobiont functioning? Trends Microbiol. 23, 490–497 (2015). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials