Motility and phototaxis of Gonium, the simplest differentiated colonial alga
- PMID: 32168596
- PMCID: PMC7616084
- DOI: 10.1103/PhysRevE.101.022416
Motility and phototaxis of Gonium, the simplest differentiated colonial alga
Abstract
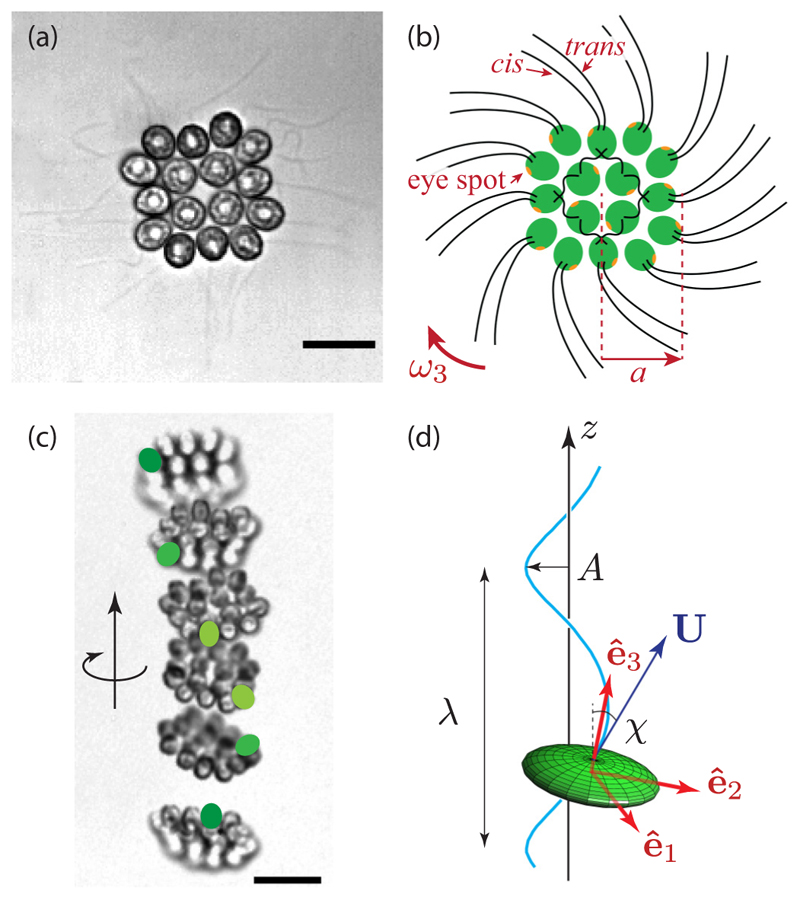
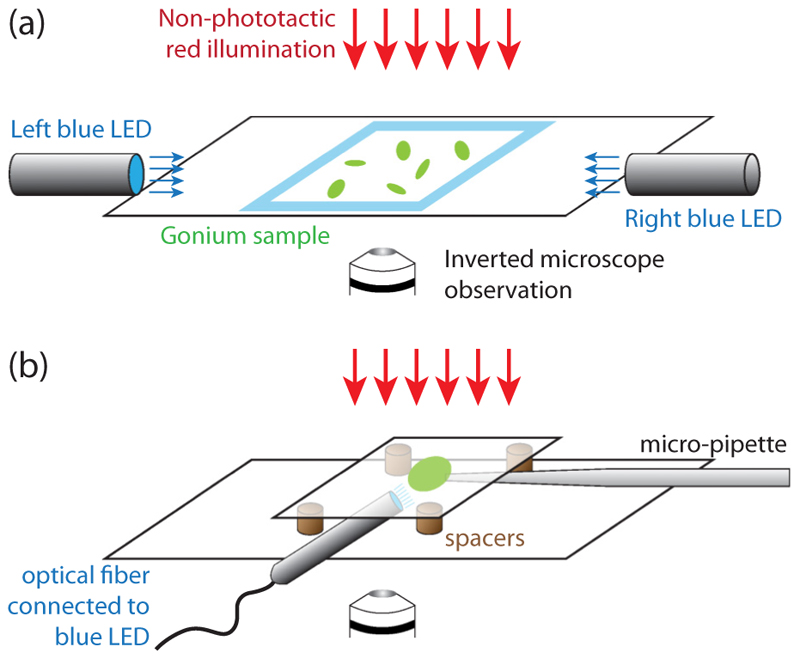
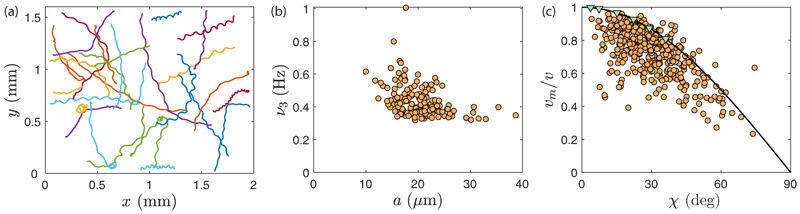
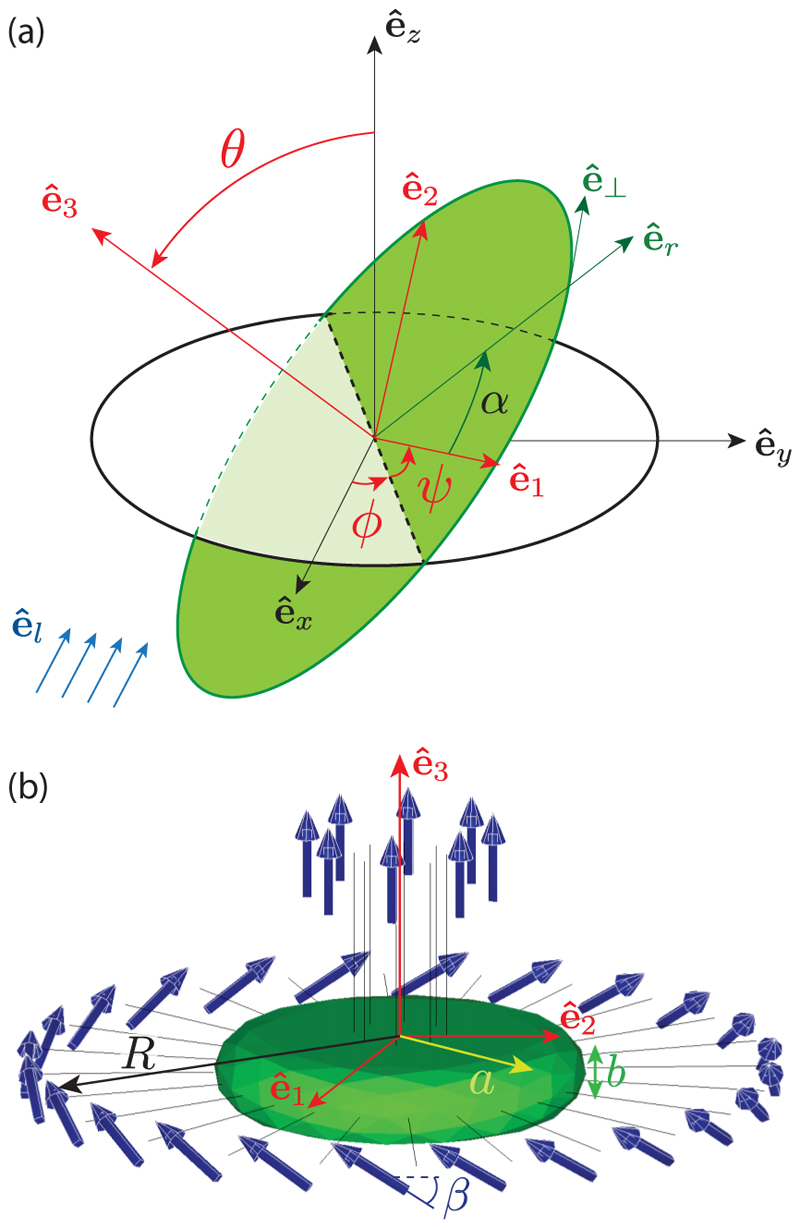
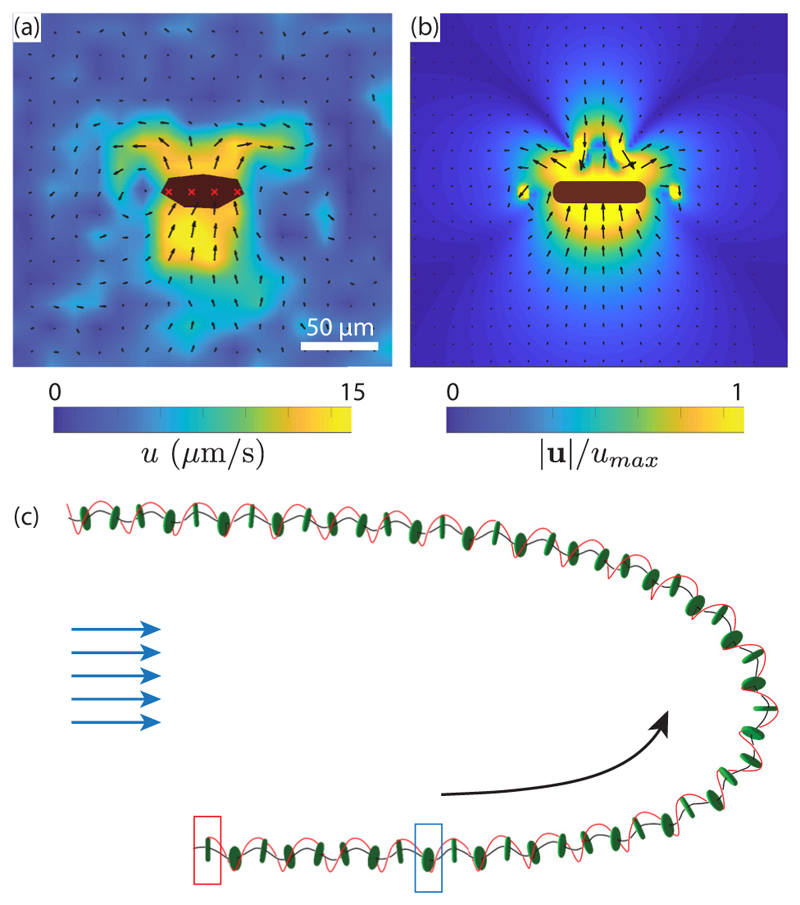
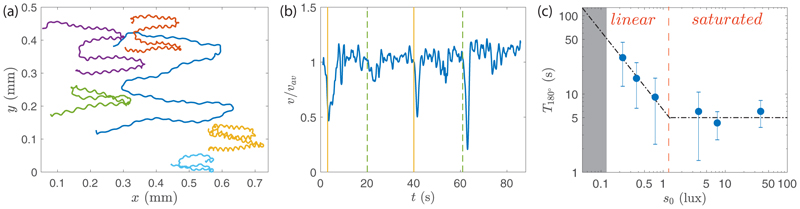
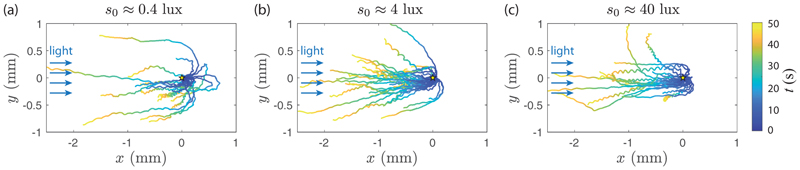
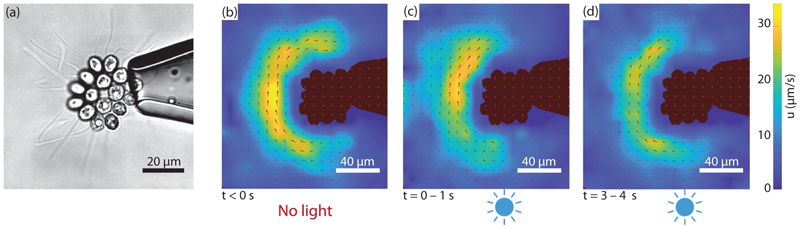
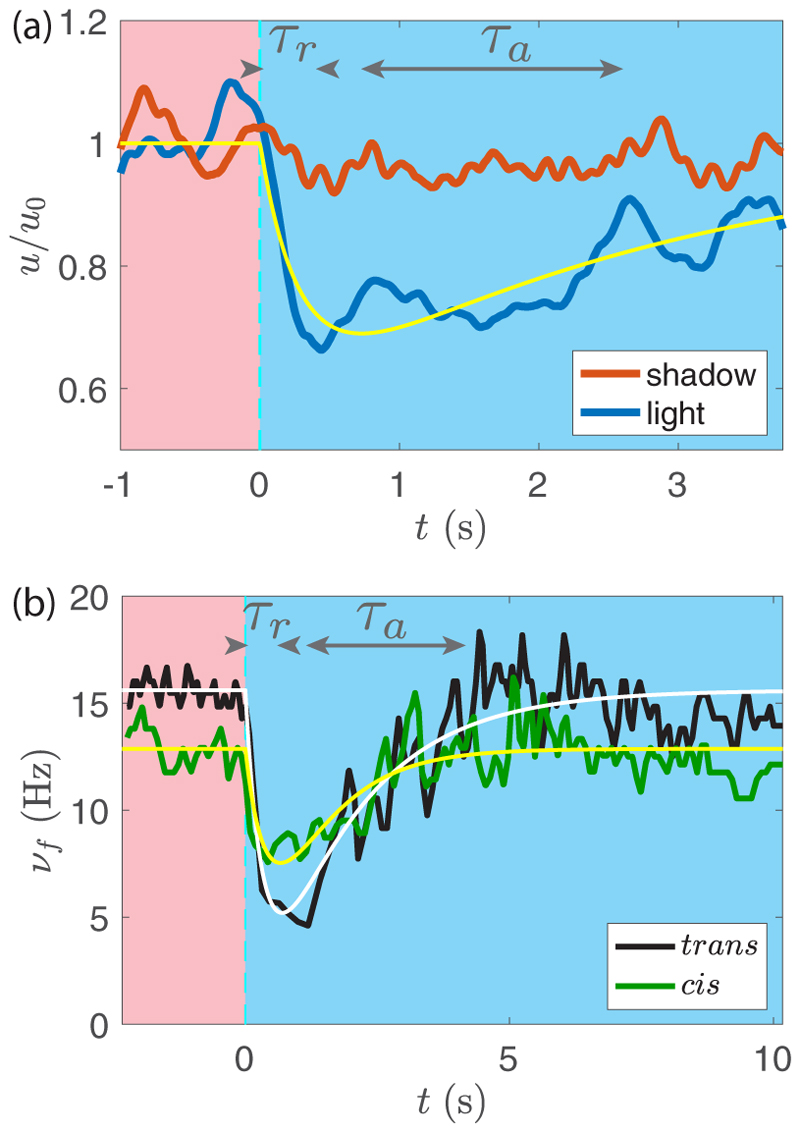
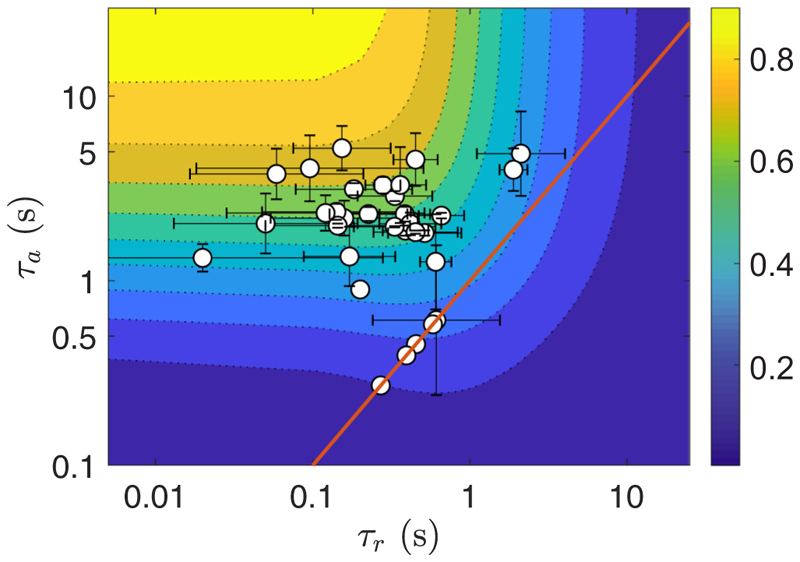
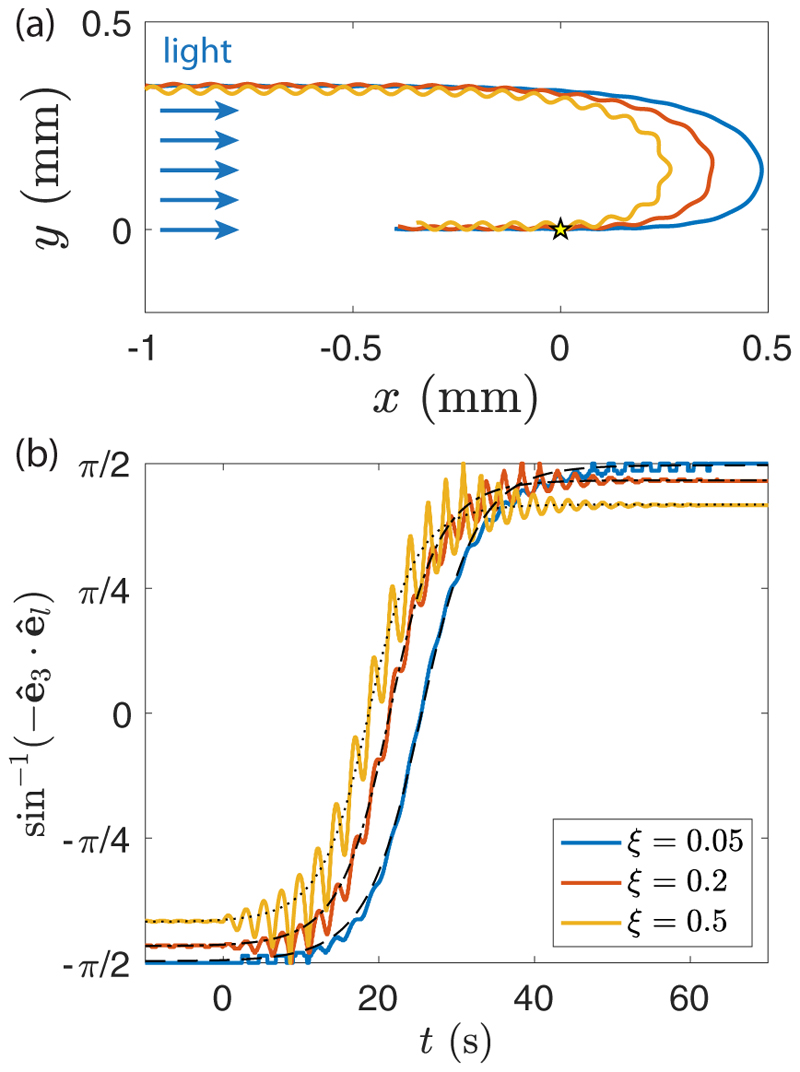
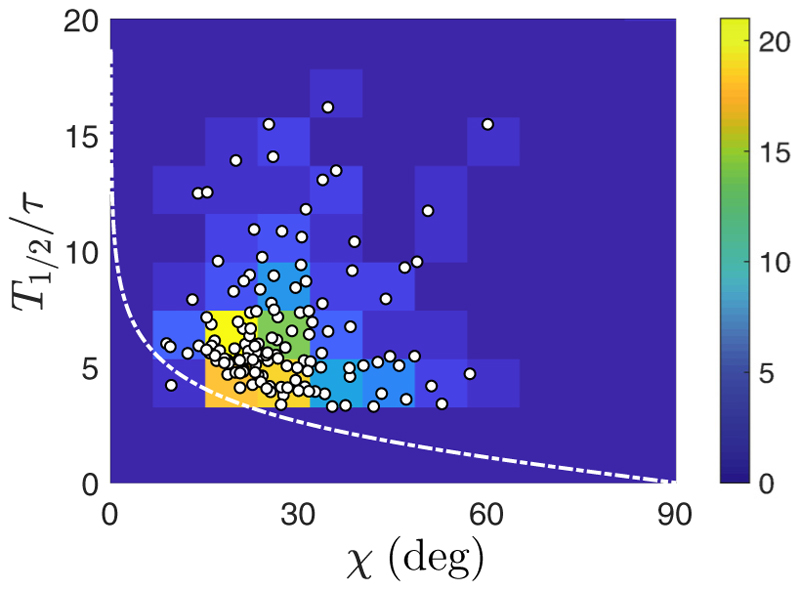
Green algae of the Volvocine lineage, spanning from unicellular Chlamydomonas to vastly larger Volvox, are models for the study of the evolution of multicellularity, flagellar dynamics, and developmental processes. Phototactic steering in these organisms occurs without a central nervous system, driven solely by the response of individual cells. All such algae spin about a body-fixed axis as they swim; directional photosensors on each cell thus receive periodic signals when that axis is not aligned with the light. The flagella of Chlamydomonas and Volvox both exhibit an adaptive response to such signals in a manner that allows for accurate phototaxis, but in the former the two flagella have distinct responses, while the thousands of flagella on the surface of spherical Volvox colonies have essentially identical behavior. The planar 16-cell species Gonium pectorale thus presents a conundrum, for its central 4 cells have a Chlamydomonas-like beat that provide propulsion normal to the plane, while its 12 peripheral cells generate rotation around the normal through a Volvox-like beat. Here we combine experiment, theory, and computations to reveal how Gonium, perhaps the simplest differentiated colonial organism, achieves phototaxis. High-resolution cell tracking, particle image velocimetry of flagellar driven flows, and high-speed imaging of flagella on micropipette-held colonies show how, in the context of a recently introduced model for Chlamydomonas phototaxis, an adaptive response of the peripheral cells alone leads to photoreorientation of the entire colony. The analysis also highlights the importance of local variations in flagellar beat dynamics within a given colony, which can lead to enhanced reorientation dynamics.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Weismann A. Essays on Heredity and Kindred Biological Problems. Clarendon Press; Oxford, UK: 1892.
-
- Huxley J. The Individual in the Animal Kingdom. Cambridge University Press; Cambridge, UK: 1912.
-
- Kirk D. Volvox Molecular-Genetic Origins of Multicellularity and Cellular Differentiation. Cambridge University Press; Cambridge, UK: 1998.
-
- Goldstein RE. Batchelor prize lecture. fluid dynamics at the scale of the cell. J Fluid Mech. 2016;807:1.
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources