Culture-independent and culture-dependent analyses of the bacterial community in the phycosphere of cyanobloom-forming Microcystis aeruginosa
- PMID: 31892695
- PMCID: PMC6938486
- DOI: 10.1038/s41598-019-56882-1
Culture-independent and culture-dependent analyses of the bacterial community in the phycosphere of cyanobloom-forming Microcystis aeruginosa
Abstract
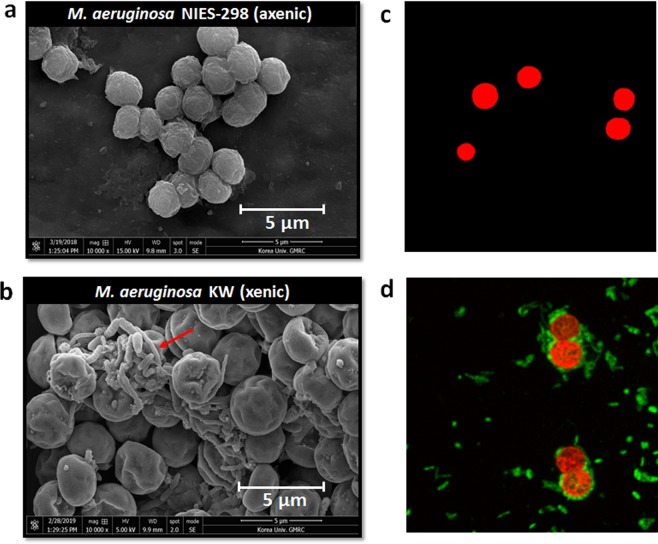
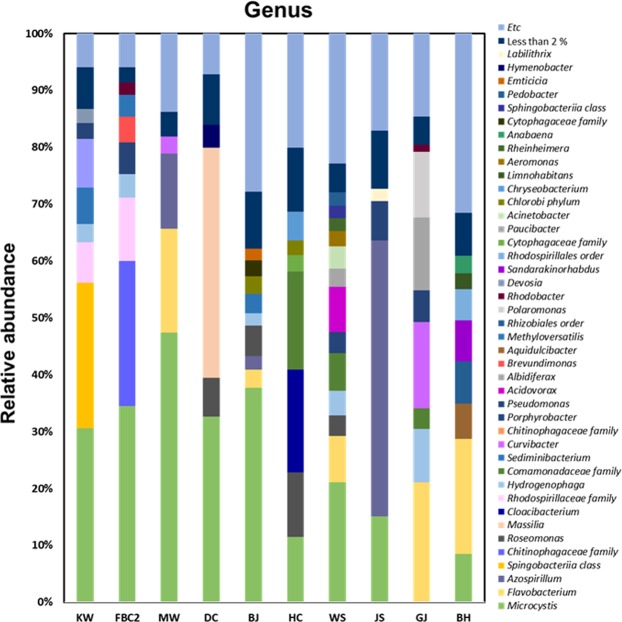
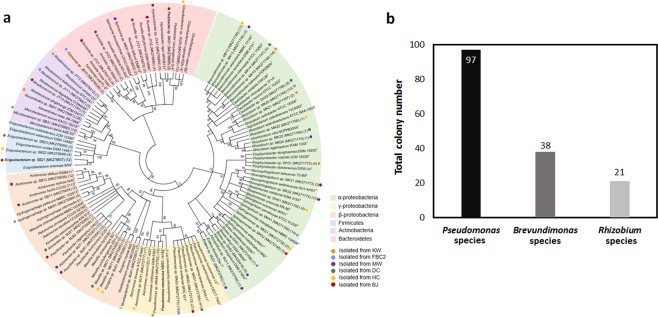
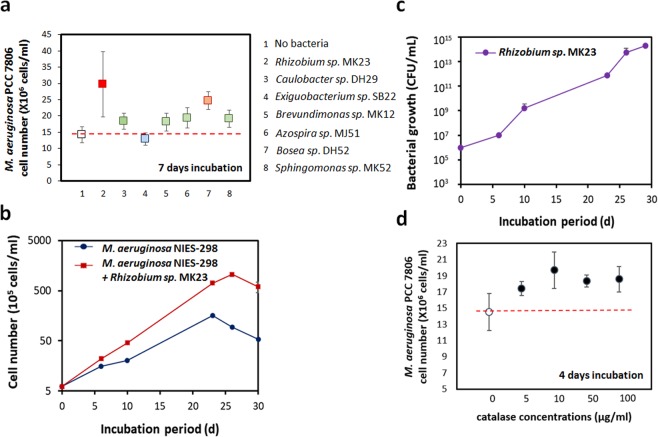
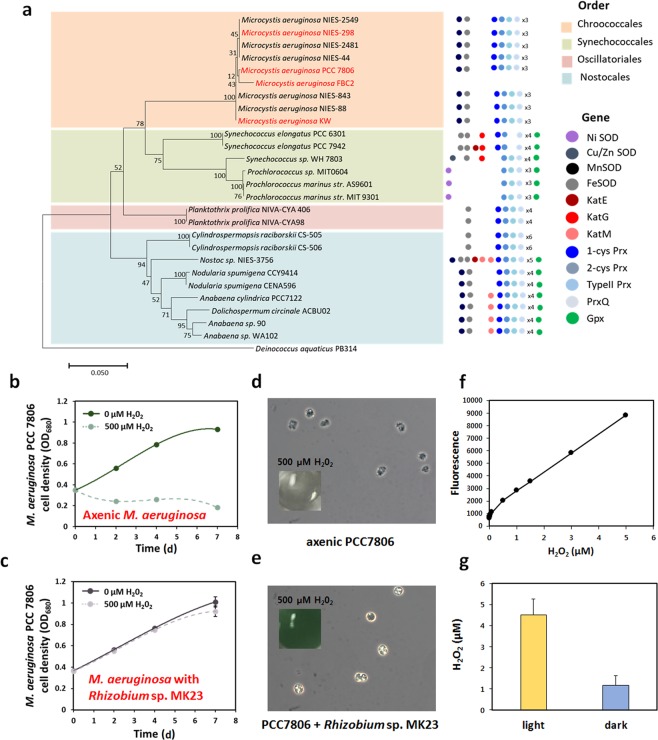
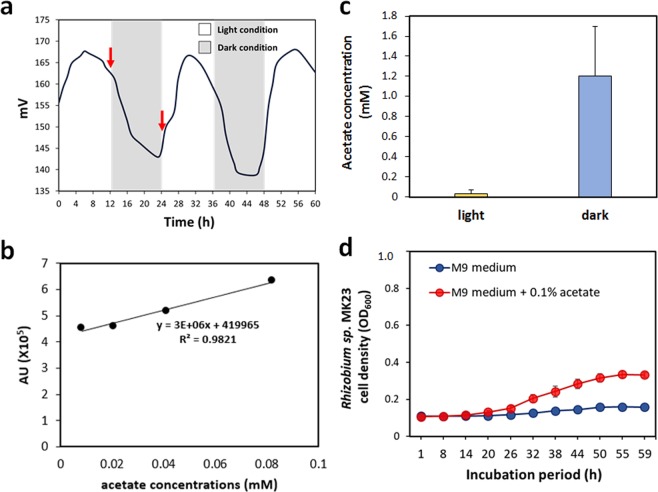
Confocal and scanning electron microscopic observations have previously shown the strong bacterial association of Microcystis aeruginosa cells on their surfaces. DNA-based analyses of the associated bacterial communities were carried out using two M. aeruginosa strains grown in the laboratory and eight newly collected cyanobacterial bloom samples. M. aeruginosa was the most predominant species (66-100%) within the phylum Cyanobacteria. Rhizobium, Hydrogenophaga and Brevundimonas species were commonly found, and Flavobacterium species were present in all the cyanobacterial bloom samples. In total, 396 colonies from various samples were screened, revealing that most culturable bacteria belonged to the class Alphaproteobacteria (19%) including Rhizobium, Brevundimonas, and Porphyrobacter species. The genetic variation among the M. aeruginosa strains and different habitat conditions may have led to the presence of distinct bacterial populations among the tested samples. Among all the tested seven culturable isolates, Rhizobium sp. MK23 showed the best growth-promotion effect on the axenic M. aeruginosa strains. H2O2 was observed to be produced during the growth of M. aeruginosa PCC7806 under light conditions, this strain was more resistant to H2O2 when associated with Rhizobium sp. MK23. Our data suggested that Rhizobium species along with other associated bacteria might help the growth of M. aeruginosa by decomposing H2O2 under the aerobic growing conditions.
Conflict of interest statement
The authors declare no competing interests.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Rabalais NN, et al. Dynamics and distribution of natural and human-caused hypoxia. Biogeosciences. 2010;7:585–619. doi: 10.5194/bg-7-585-2010. - DOI
-
- Lewis WM, Wurtsbaugh WA. Control of lacustrine phytoplankton by nutrients: erosion of the phosphorus paradigm. Intern. Rev. Hydrobiol. 2008;93:446–465. doi: 10.1002/iroh.200811065. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials