Extracellular polysaccharide synthesis in a bloom-forming strain of Microcystis aeruginosa: implications for colonization and buoyancy
- PMID: 30718739
- PMCID: PMC6362013
- DOI: 10.1038/s41598-018-37398-6
Extracellular polysaccharide synthesis in a bloom-forming strain of Microcystis aeruginosa: implications for colonization and buoyancy
Abstract
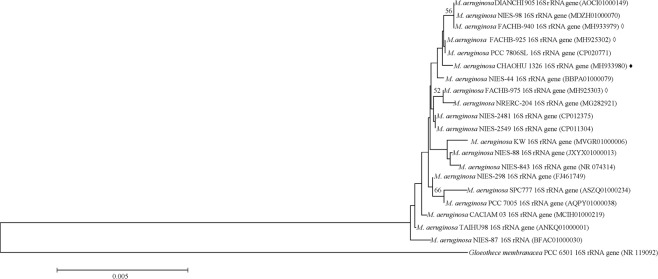
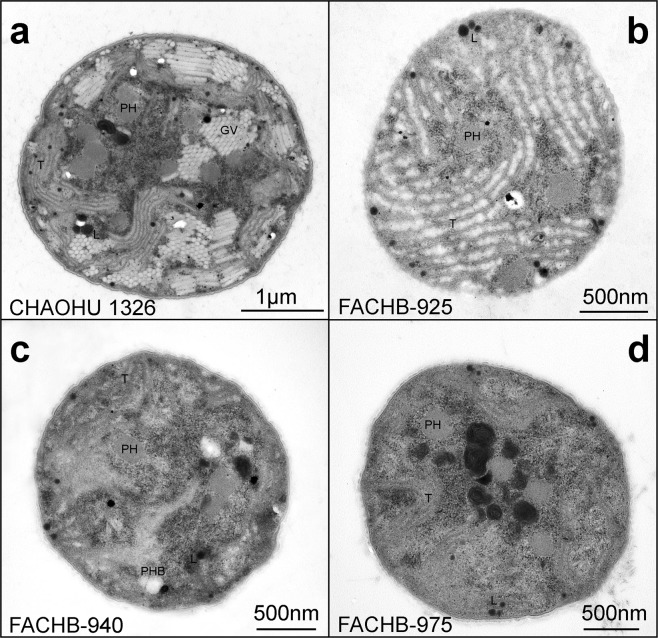
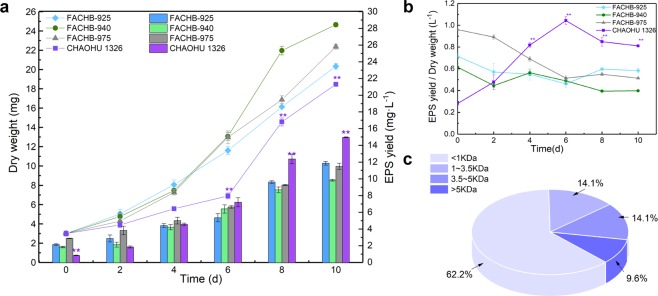
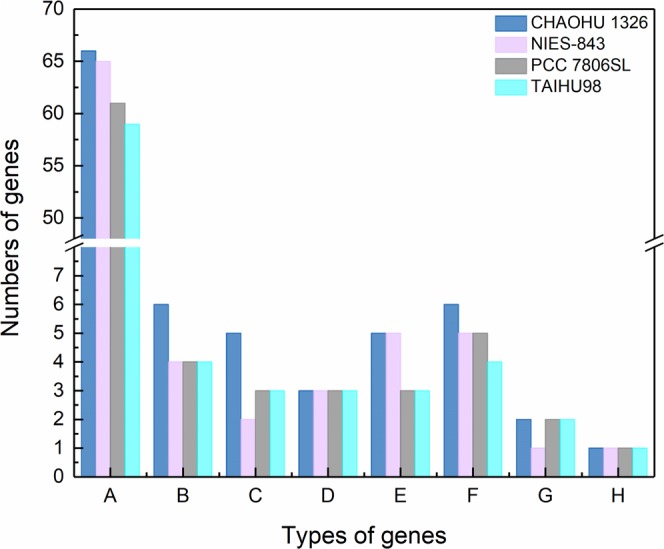
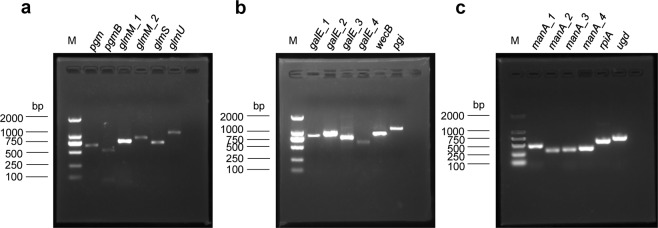
Microcystis, the dominant species among cyanobacterial blooms, normally forms colonies under natural conditions but exists as single cells or paired cells in axenic laboratory cultures after long-term cultivation. Here, a bloom-forming Microcystis aeruginosa strain CHAOHU 1326 was studied because it presents a colonial morphology and grows on the water surface during axenic laboratory culturing. We first examined the morphological features of strain CHAOHU 1326 and three other unicellular M. aeruginosa strains FACHB-925, FACHB-940, and FACHB-975 cultured under the same conditions by scanning and transmission electron microscopy. Then, we compared the extracellular polysaccharide (EPS)-producing ability of colonial strain CHAOHU 1326 to that of the three unicellular M. aeruginosa strains, and found that strain CHAOHU 1326 produced a higher amount of EPS than the other strains during growth. Moreover, based on genome sequencing, multiple gene clusters implicated in EPS biosynthesis and a cluster of 12 genes predicted to be involved in gas vesicle synthesis in strain CHAOHU 1326 were detected. These predicted genes were all functional and expressed in M. aeruginosa CHAOHU 1326 as determined by reverse transcription PCR. These findings provide a physiological and genetic basis to better understand colony formation and buoyancy control during M. aeruginosa blooming.
Conflict of interest statement
The authors declare no competing interests.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Šejnohová, L. & Maršálek, B. In Ecology of Cyanobacteria II: Their Diversity in Space and Time (ed. Brian, A. W.) 195–228 (Springer Netherlands, 2012).
-
- Codd GA, Lindsay J, Young FM, Morrison LF, Metcalf JS. Harmful cyanobacteria. Aquatic Ecology. 2005;3:1–23. doi: 10.1007/1-4020-3022-3_1. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials