Repeated evolution and reversibility of self-fertilization in the volvocine green algae
- PMID: 29134623
- PMCID: PMC5796843
- DOI: 10.1111/evo.13394
Repeated evolution and reversibility of self-fertilization in the volvocine green algae
Abstract
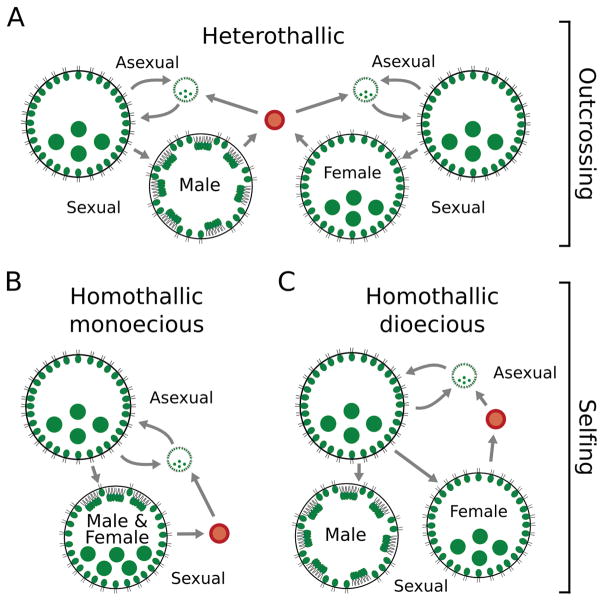
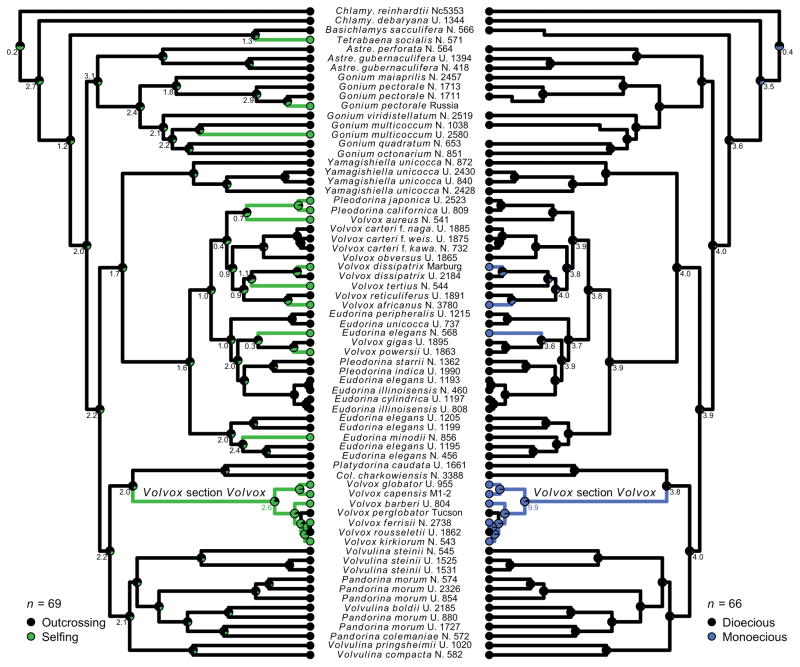
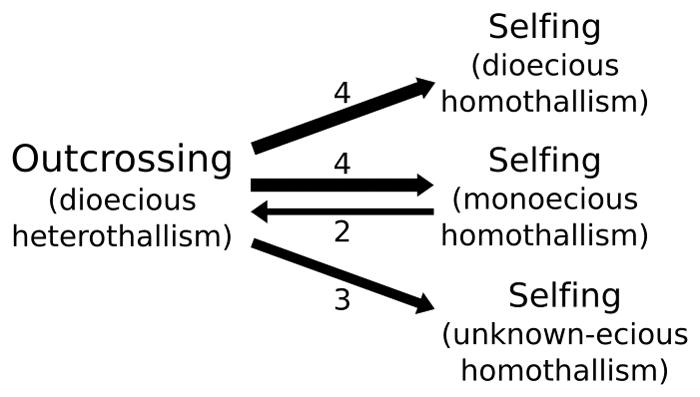
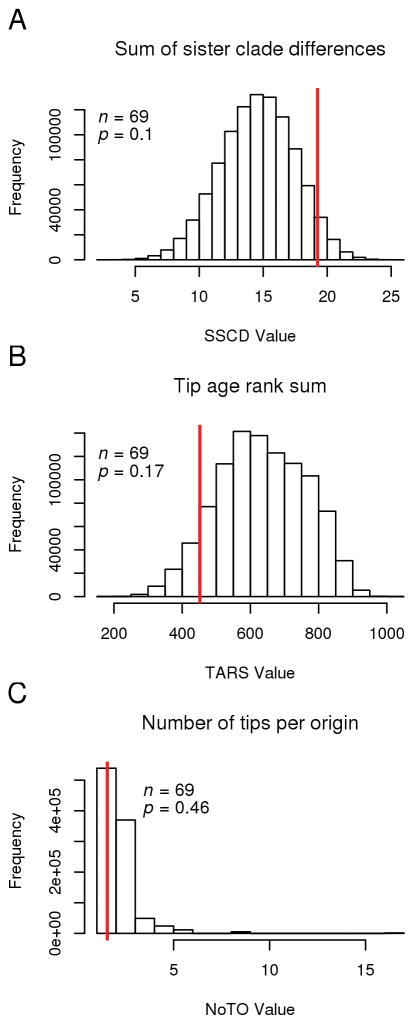
Outcrossing and self-fertilization are fundamental strategies of sexual reproduction, each with different evolutionary costs and benefits. Self-fertilization is thought to be an evolutionary "dead-end" strategy, beneficial in the short term but costly in the long term, resulting in self-fertilizing species that occupy only the tips of phylogenetic trees. Here, we use volvocine green algae to investigate the evolution of self-fertilization. We use ancestral-state reconstructions to show that self-fertilization has repeatedly evolved from outcrossing ancestors and that multiple reversals from selfing to outcrossing have occurred. We use three phylogenetic metrics to show that self-fertilization is not restricted to the tips of the phylogenetic tree, a finding inconsistent with the view of self-fertilization as a dead-end strategy. We also find no evidence for higher extinction rates or lower speciation rates in selfing lineages. We find that self-fertilizing species have significantly larger colonies than outcrossing species, suggesting the benefits of selfing may counteract the costs of increased size. We speculate that our macroevolutionary results on self-fertilization (i.e., non-tippy distribution, no decreased diversification rates) may be explained by the haploid-dominant life cycle that occurs in volvocine algae, which may alter the costs and benefits of selfing.
Keywords: Haploid; phylogenetics; self-fertilization; sex; sexual reproduction; volvocine green algae.
© 2017 The Author(s). Evolution © 2017 The Society for the Study of Evolution.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Comment in
-
Digest: Going solo: Self-fertilization in haploid algae may not lead to evolutionary decline.Vincent AM. Vincent AM. Evolution. 2018 Feb;72(2):409-410. doi: 10.1111/evo.13420. Evolution. 2018. PMID: 29315523 No abstract available.
References
-
- Akaike H. A new look at the statistical model identification. IEEE Trans Automat Contr. 1974;19:716–723.
-
- Baker HG. Self-compatibility and establishment after "long-distance" dispersal. Evolution. 1955;9:347–349.
-
- Barrett SCH. The evolution of plant sexual diversity. Nat Rev Genet. 2002;3:274–284. - PubMed
-
- Bawa KS. Evolution of dioecy in flowering plants. Annu Rev Ecol Syst. 1980;11:15–39.
Publication types
MeSH terms
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials