A new paradigm for producing astaxanthin from the unicellular green alga Haematococcus pluvialis
- PMID: 27563850
- PMCID: PMC5071682
- DOI: 10.1002/bit.25976
A new paradigm for producing astaxanthin from the unicellular green alga Haematococcus pluvialis
Abstract
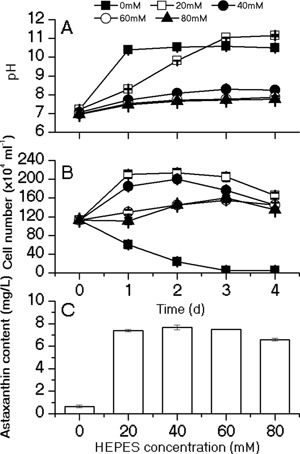
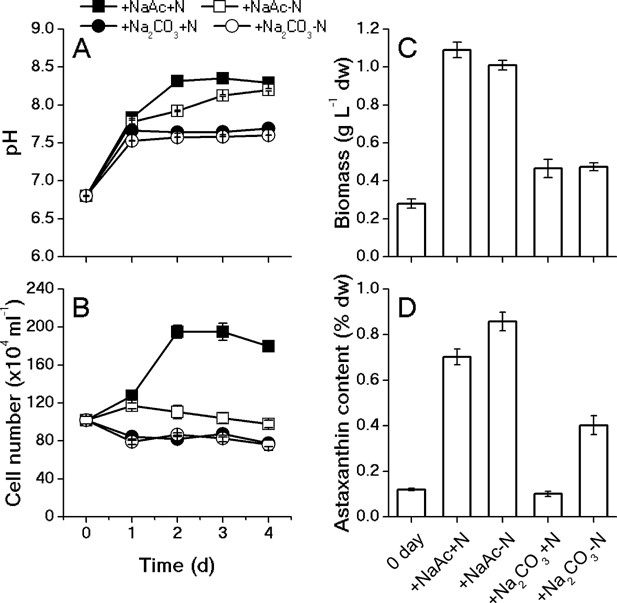
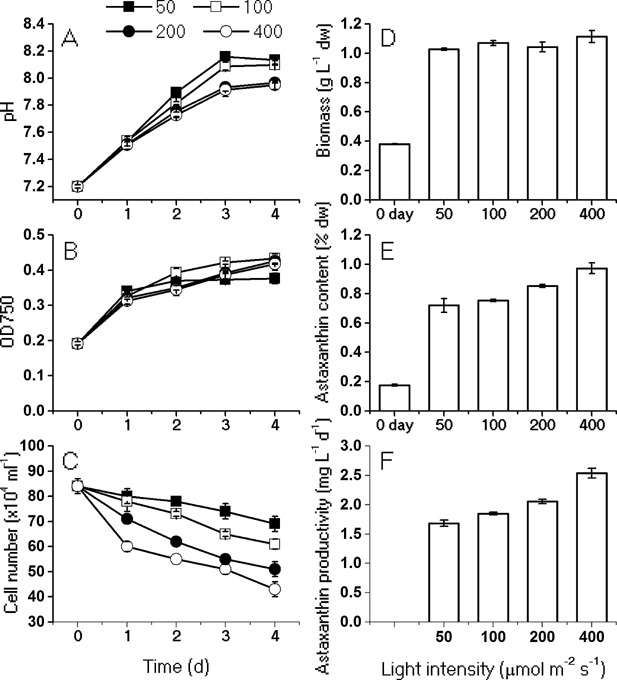
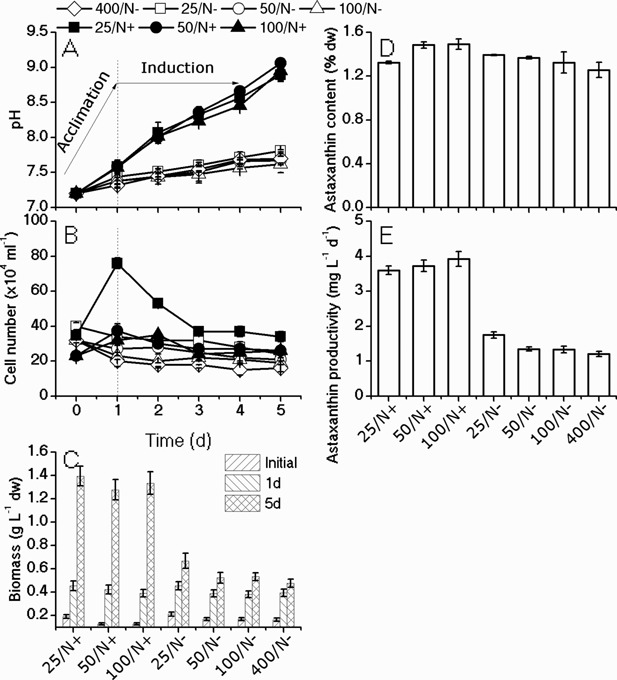
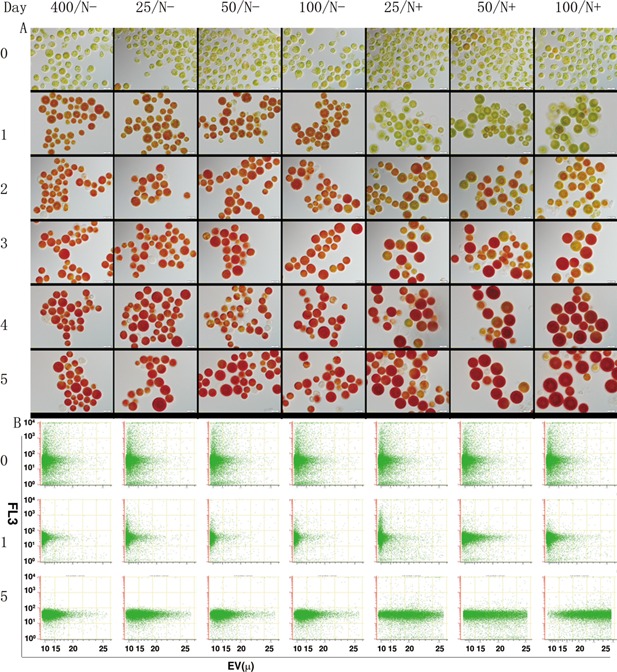
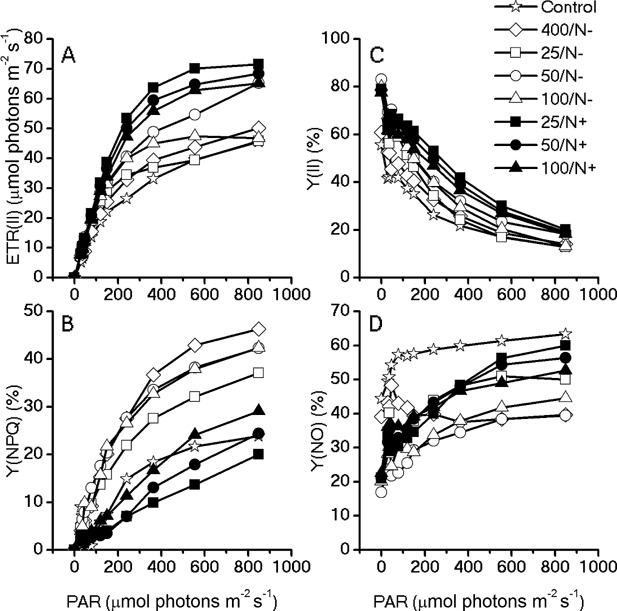
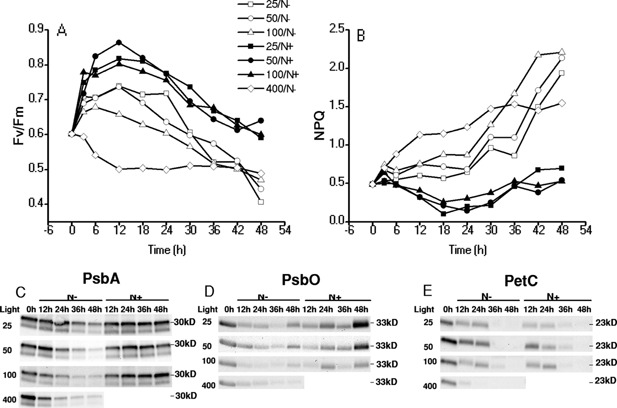
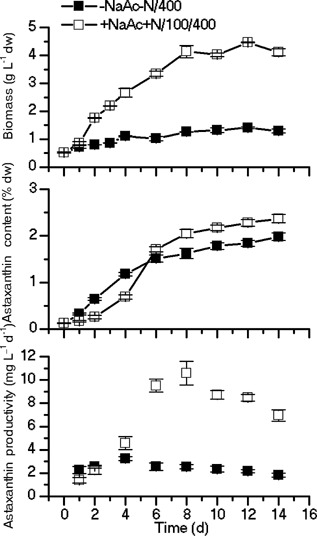
The unicellular green alga Haematococcus pluvialis has been exploited as a cell factory to produce the high-value antioxidant astaxanthin for over two decades, due to its superior ability to synthesize astaxanthin under adverse culture conditions. However, slow vegetative growth under favorable culture conditions and cell deterioration or death under stress conditions (e.g., high light, nitrogen starvation) has limited the astaxanthin production. In this study, a new paradigm that integrated heterotrophic cultivation, acclimation of heterotrophically grown cells to specific light/nutrient regimes, followed by induction of astaxanthin accumulation under photoautotrophic conditions was developed. First, the environmental conditions such as pH, carbon source, nitrogen regime, and light intensity, were optimized to induce astaxanthin accumulation in the dark-grown cells. Although moderate astaxanthin content (e.g., 1% of dry weight) and astaxanthin productivity (2.5 mg L(-1) day(-1) ) were obtained under the optimized conditions, a considerable number of cells died off when subjected to stress for astaxanthin induction. To minimize the susceptibility of dark-grown cells to light stress, the algal cells were acclimated, prior to light induction of astaxanthin biosynthesis, under moderate illumination in the presence of nitrogen. Introduction of this strategy significantly reduced the cell mortality rate under high-light and resulted in increased cellular astaxanthin content and astaxanthin productivity. The productivity of astaxanthin was further improved to 10.5 mg L(-1) day(-1) by implementation of such a strategy in a bubbling column photobioreactor. Biochemical and physiological analyses suggested that rebuilding of photosynthetic apparatus including D1 protein and PsbO, and recovery of PSII activities, are essential for acclimation of dark-grown cells under photo-induction conditions. Biotechnol. Bioeng. 2016;113: 2088-2099. © 2016 The Authors. Biotechnology and Bioengineering Published by Wiley Periodicals, Inc.
Keywords: Haematococcus pluvialis; acclimation; astaxanthin; heterotrophy.
© 2016 The Authors. Biotechnology and Bioengineering Published by Wiley Periodicals, Inc.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Aflalo C, Meshulam Y, Zarka A, Boussiba S. 2007. On the relative efficiency of two‐vs. one‐stage production of astaxanthin by the green alga Haematococcus pluvialis . Biotechnol Bioeng 98(1):300–305. - PubMed
-
- Baker NR. 2008. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu Rev Plant Biol 59:89–113. - PubMed
-
- Björkman O, Demmig B. 1987. Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77K among vascular plants of diverse origins. Planta 170(4):489–504. - PubMed
-
- Boussiba S, Vonshak A. 1991. Astaxanthin accumulation in the green alga Haematococcus pluvialis . Plant Cell Physiol 32(7):1077–1082.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials