Immortalization of Fetal Bovine Colon Epithelial Cells by Expression of Human Cyclin D1, Mutant Cyclin Dependent Kinase 4, and Telomerase Reverse Transcriptase: An In Vitro Model for Bacterial Infection
- PMID: 26624883
- PMCID: PMC4666463
- DOI: 10.1371/journal.pone.0143473
Immortalization of Fetal Bovine Colon Epithelial Cells by Expression of Human Cyclin D1, Mutant Cyclin Dependent Kinase 4, and Telomerase Reverse Transcriptase: An In Vitro Model for Bacterial Infection
Abstract
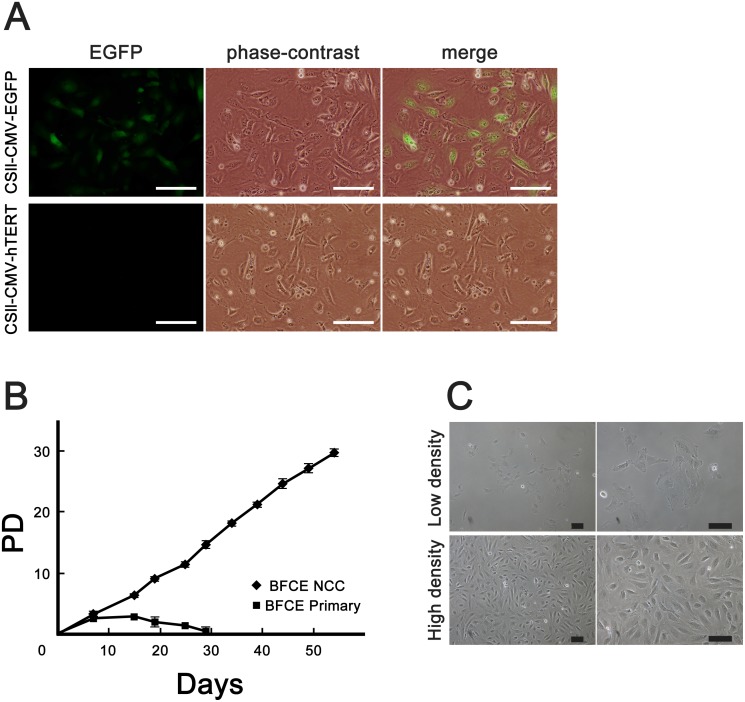
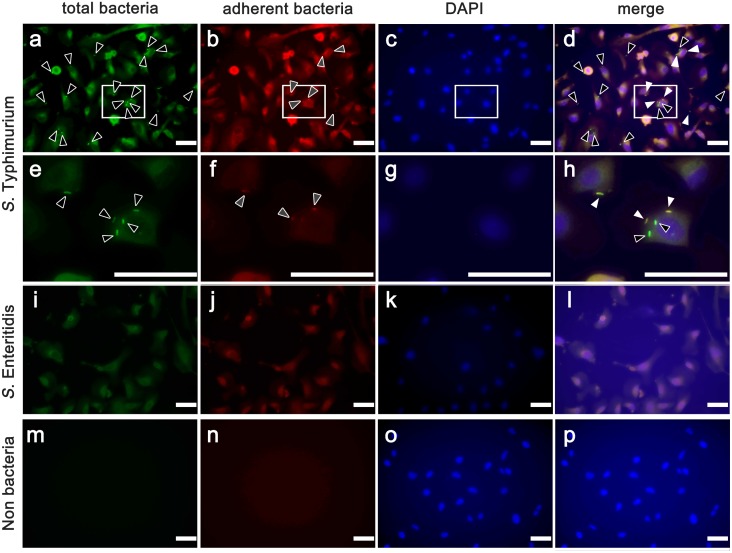
Cattle are the economically important animals in human society. They are essential for the production of livestock products such as milk and meats. The production efficiency of livestock products is negatively impacted by infection with zoonotic pathogens. To prevent and control infectious diseases, it is important to understand the interaction between cattle tissue and pathogenic bacteria. In this study, we established an in vitro infection model of an immortalized bovine colon-derived epithelial cell line by transducing the cells with lentiviral vectors containing genes encoding cell cycle regulators cyclin D1, mutant cyclin dependent kinase 4 (CDK4), and human telomerase reverse transcriptase (TERT). The established cell line showed continuous cell proliferation, expression of epithelial markers, and an intact karyotype, indicating that the cells maintained their original nature as colon-derived epithelium. Furthermore, we exposed the established cell line to two strains of Salmonella enterica and EHEC. Interestingly, S. Typhimurium showed higher affinity for the established cell line and invaded the cytoplasm than S. Enteritidis. Quantitative RT-PCR revealed that gene expression of Toll-like receptor 1 (TLR1), TLR 2 and TLR 3, whereas TLR 4, 5 and 6 were not detectable in established cells. Our established immortalized colon-derived epithelial cell should be a useful tool for studies evaluating the molecular mechanisms underlying bacterial infection.
Conflict of interest statement
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Rosselin M, Abed N, Virlogeux-Payant I, Bottreau E, Sizaret PY, Velge P, et al. Heterogeneity of type III secretion system (T3SS)-1-independent entry mechanisms used by Salmonella Enteritidis to invade different cell types. Microbiology. 2011;157(Pt 3):839–47. Epub 2010年11月27日. 10.1099/mic.0.044941-0 . - DOI - PubMed
-
- Ozer HL, Banga SS, Dasgupta T, Houghton J, Hubbard K, Jha KK, et al. SV40-mediated immortalization of human fibroblasts. Exp Gerontol. 1996;31(1–2):303–10. Epub 1996年01月01日. . - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials