Delineating a New Heterothallic Species of Volvox (Volvocaceae, Chlorophyceae) Using New Strains of "Volvox africanus"
- PMID: 26562165
- PMCID: PMC4643018
- DOI: 10.1371/journal.pone.0142632
Delineating a New Heterothallic Species of Volvox (Volvocaceae, Chlorophyceae) Using New Strains of "Volvox africanus"
Abstract
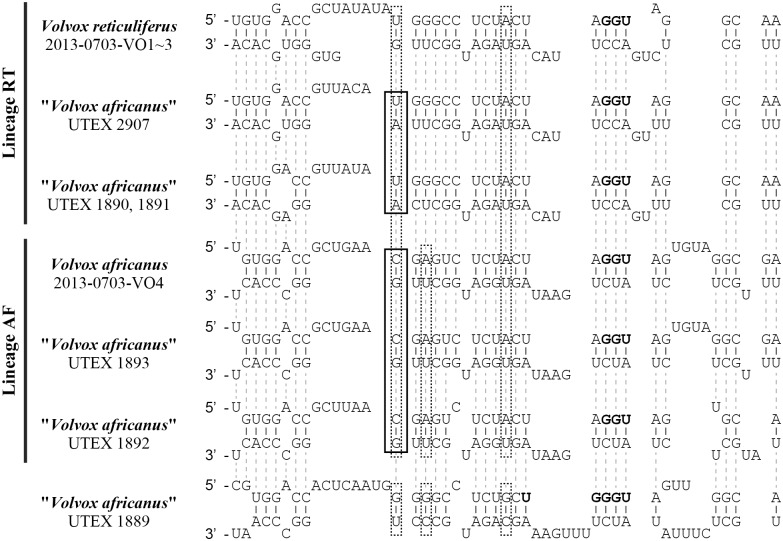
The volvocine algae represent an excellent model lineage in which to study evolution of female and male genders based on comparative analyses of related species. Among these species, Volvox carteri has been extensively studied as a model of an oogamous and complex organism. However, it may have unique derived features that are not present in other species of Volvox. Therefore, information regarding the characteristics of sexual reproduction of other species of Volvox is also important. In 1971, Starr studied four types of sexuality in several global strains identified as Volvox africanus; however, further taxonomic studies of these strains have been lacking, and strains of three of the four sexual types are not available. Here, we studied the morphology, sexual reproduction, and taxonomy of two V. africanus-like species isolated recently from Lake Biwa, Japan. These two species were very similar to two sexual types described by Starr in 1971: one producing dioecious sexual spheroids in heterothallic strains and the other forming both male spheroids and monoecious spheroids in a single strain. The former species produced zygotes with a reticulate cell wall, whereas a smooth zygote wall was observed in the latter species as in V. africanus previously reported from various localities around the world. Our multigene phylogenetic analysis demonstrated that these are sister species to each other. However, the presence of a compensatory base change in the most conserved region of the secondary structure of nuclear ribosomal DNA internal transcribed spacer-2, hybrid inviability demonstrated by intercrossing experiments, and morphological differences in the density of abutment between the gelatinous material of adjacent cells (individual sheaths) in the spheroid supported the recognition of the two species, V. africanus having a smooth zygote wall and V. reticuliferus Nozaki sp. nov. having a reticulate zygote wall.
Conflict of interest statement
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Linnaeus C. Systema naturae Regnum animalae. Ed. 10, Stockholm; 1758.
-
- Smith GM. A comparative study of the species of Volvox Trans. Am. Microsc. Soc. 1944; 63: 265–310.
-
- Nozaki H. Morphology, sexual reproduction and taxonomy of Volvox carteri f. kawasakiensis f. nov. (Chlorophyta) from Japan Phycologia 1988; 27: 209–220.
-
- Kirk DL. Volvox: Molecular Genetic Origins of Multicellularity and Cellular Differentiation. Cambridge: Cambridge University Press; 1998.
-
- Nozaki H, Coleman AW. A new species of Volvox sect. Merrillosphaera (Volvocaceae, Chlorophyceae) from Texas. J. Phycol. 2011;47: 673–679. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials