Extensive horizontal gene transfer, duplication, and loss of chlorophyll synthesis genes in the algae
- PMID: 25887237
- PMCID: PMC4337275
- DOI: 10.1186/s12862-015-0286-4
Extensive horizontal gene transfer, duplication, and loss of chlorophyll synthesis genes in the algae
Abstract
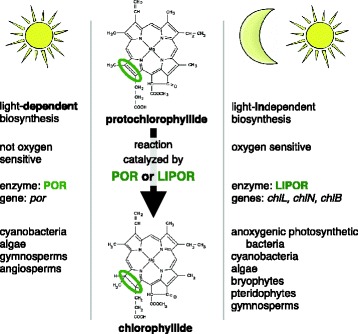
Background: Two non-homologous, isofunctional enzymes catalyze the penultimate step of chlorophyll a synthesis in oxygenic photosynthetic organisms such as cyanobacteria, eukaryotic algae and land plants: the light-independent (LIPOR) and light-dependent (POR) protochlorophyllide oxidoreductases. Whereas the distribution of these enzymes in cyanobacteria and land plants is well understood, the presence, loss, duplication, and replacement of these genes have not been surveyed in the polyphyletic and remarkably diverse eukaryotic algal lineages.
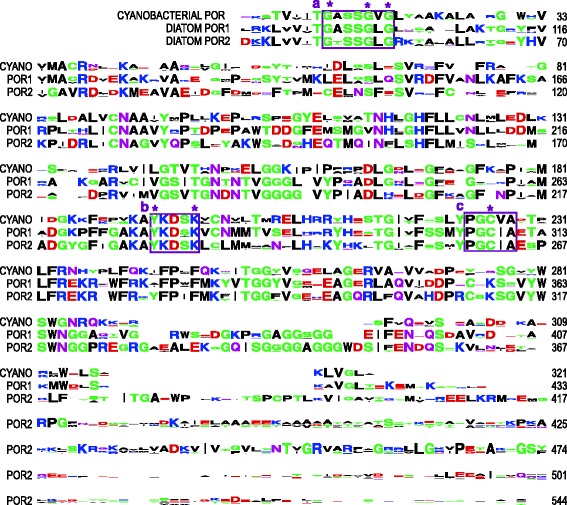
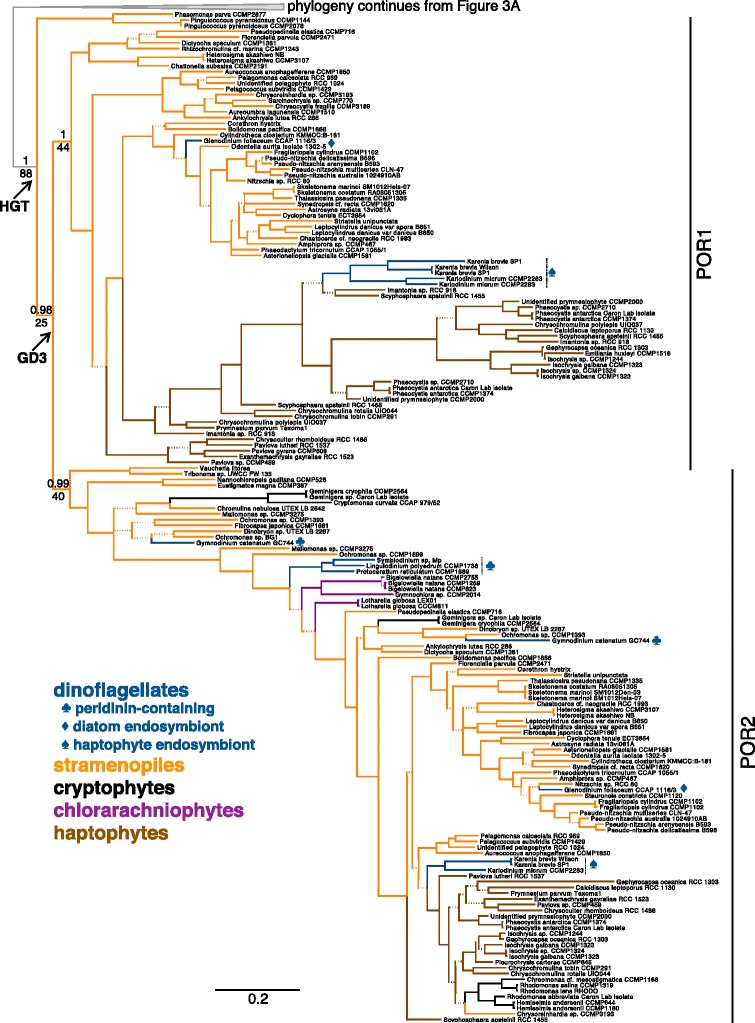
Results: A phylogenetic reconstruction of the history of the POR enzyme (encoded by the por gene in nuclei) in eukaryotic algae reveals replacement and supplementation of ancestral por genes in several taxa with horizontally transferred por genes from other eukaryotic algae. For example, stramenopiles and haptophytes share por gene duplicates of prasinophytic origin, although their plastid ancestry predicts a rhodophytic por signal. Phylogenetically, stramenopile pors appear ancestral to those found in haptophytes, suggesting transfer from stramenopiles to haptophytes by either horizontal or endosymbiotic gene transfer. In dinoflagellates whose plastids have been replaced by those of a haptophyte or diatom, the ancestral por genes seem to have been lost whereas those of the new symbiotic partner are present. Furthermore, many chlorarachniophytes and peridinin-containing dinoflagellates possess por gene duplicates. In contrast to the retention, gain, and frequent duplication of algal por genes, the LIPOR gene complement (chloroplast-encoded chlL, chlN, and chlB genes) is often absent. LIPOR genes have been lost from haptophytes and potentially from the euglenid and chlorarachniophyte lineages. Within the chlorophytes, rhodophytes, cryptophytes, heterokonts, and chromerids, some taxa possess both POR and LIPOR genes while others lack LIPOR. The gradual process of LIPOR gene loss is evidenced in taxa possessing pseudogenes or partial LIPOR gene compliments. No horizontal transfer of LIPOR genes was detected.
Conclusions: We document a pattern of por gene acquisition and expansion as well as loss of LIPOR genes from many algal taxa, paralleling the presence of multiple por genes and lack of LIPOR genes in the angiosperms. These studies present an opportunity to compare the regulation and function of por gene families that have been acquired and expanded in patterns unique to each of various algal taxa.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Willows RD. Chlorophyll synthesis. In: Wise RR, Hoober JK, editors. The structure and function of plastids. Dordrecht: Springer; 2006. pp. 295–313.
-
- Armstrong G. Greening in the dark: light-independent chlorophyll biosynthesis from anoxygenic photosynthetic bacteria to gymnosperms. J Photochem Photobiol B Biol. 1998;43:87–100.
-
- Fujita Y, Bauer CE. The light-dependent protochlorophyllide reductase: a nitrogenase-like enzyme catalyzing a key reaction for greening in the dark. In: Kadish K, Smith K, Guilard R, editors. The porphyrin handbook. vol. 13. San Diego: Elsevier Science; 2003. pp. 109–56.
-
- Muraki N, Nomata J, Ebata K, Mizoguchi T, Shiba T, Tamiaki H, et al. X-ray crystal structure of the light-independent protochlorophyllide reductase. Nature. 2010;465:110–4. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources