Toll-like receptor 4 is a sensor for autophagy associated with innate immunity
- PMID: 17658277
- PMCID: PMC2680670
- DOI: 10.1016/j.immuni.2007年05月02日2
Toll-like receptor 4 is a sensor for autophagy associated with innate immunity
Abstract
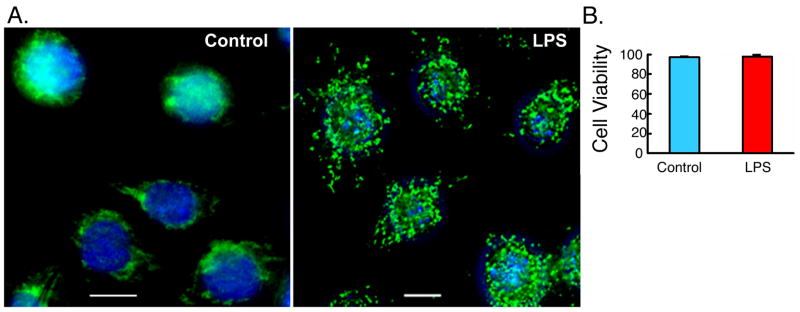
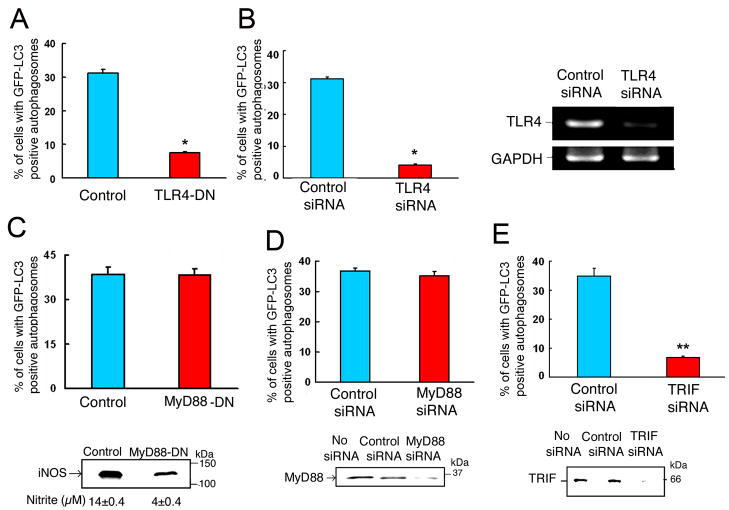
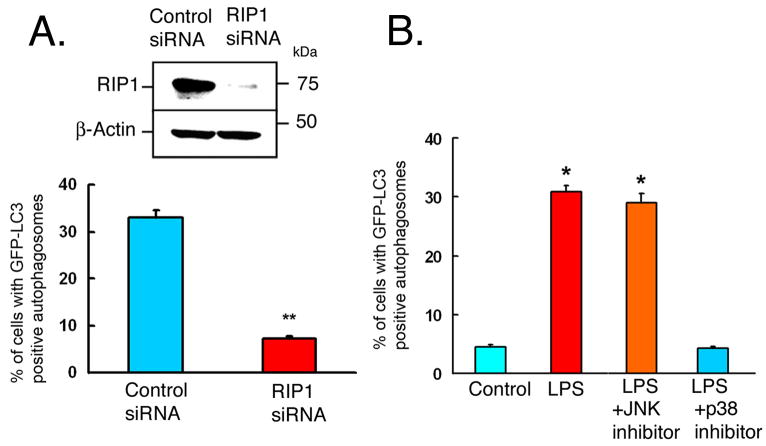
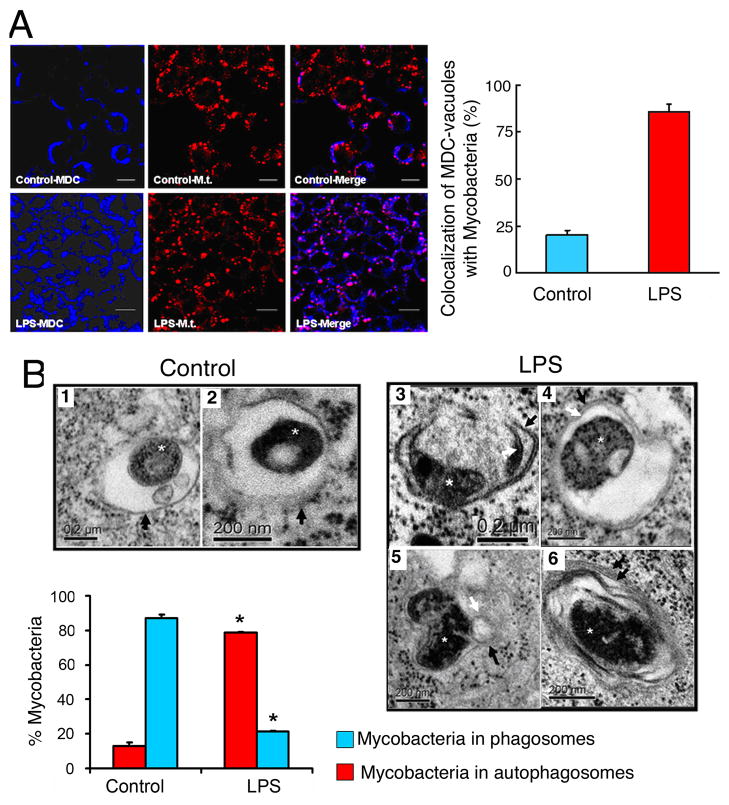
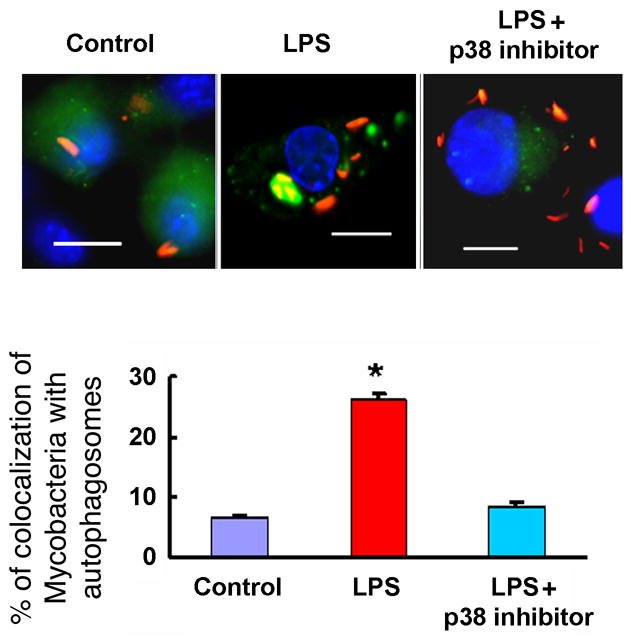
Autophagy has recently been shown to be an important component of the innate immune response. The signaling pathways leading to activation of autophagy in innate immunity are not known. Here we showed that Toll-like receptor 4 (TLR4) served as a previously unrecognized environmental sensor for autophagy. Autophagy was induced by lipopolysaccharide (LPS) in primary human macrophages and in the murine macrophage RAW264.7 cell line. We defined a new molecular pathway in which LPS-induced autophagy was regulated through a Toll-interleukin-1 receptor domain-containing adaptor-inducing interferon-beta (TRIF)-dependent, myeloid differentiation factor 88 (MyD88)-independent TLR4 signaling pathway. Receptor-interacting protein (RIP1) and p38 mitogen-activated protein kinase were downstream components of this pathway. This signaling pathway did not affect cell viability, indicating that it is distinct from the autophagic death signaling pathway. We further showed that LPS-induced autophagy could enhance mycobacterial colocalization with the autophagosomes. This study links two ancient processes, autophagy and innate immunity, together through a shared signaling pathway.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Levine B, Klionsky DJ. Development by self-digestion: molecular mechanisms and biological functions of autophagy. Dev Cell. 2004;6:463–477. - PubMed
-
- Levine B. Eating oneself and uninvited guest: autophagy-related pathways in cellular defense. Cell. 2005;120:159–162. - PubMed
-
- Nakagawa I, Amano A, Mizushima N, Yamamoto A, Yamaguchi H, Kamimoto T, Nara A, Funao J, Nakata M, Tsuda K, et al. Autophagy defends cells against invading group A Streptococcus. Science. 2004;306:1037–1040. - PubMed
-
- Ogawa M, Yoshimori T, Suzuki T, Sagara H, Mizushima N, Sasakawa C. Escape of intracellular Shigella from autophagy. Science. 2005;307:727–731. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous