fMRI spectral signatures of sleep
- PMID: 35862450
- PMCID: PMC9335231
- DOI: 10.1073/pnas.2016732119
fMRI spectral signatures of sleep
Abstract
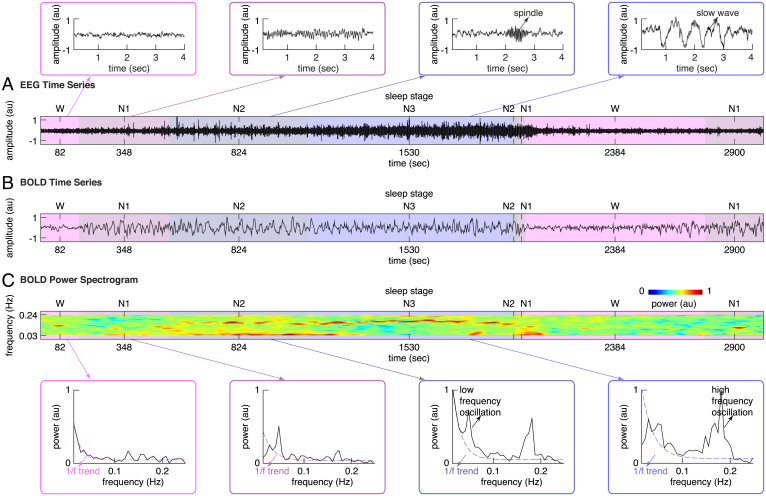
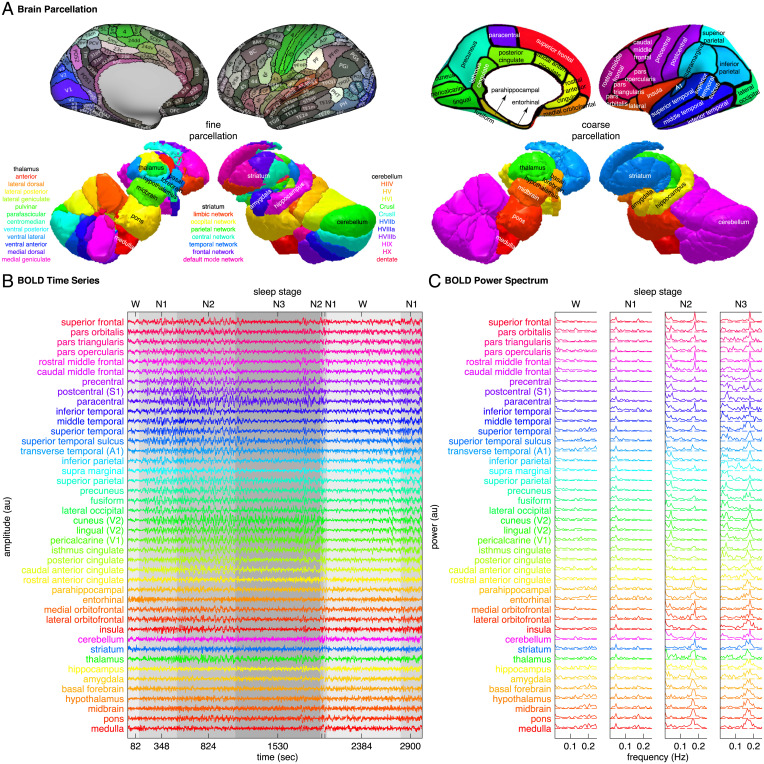
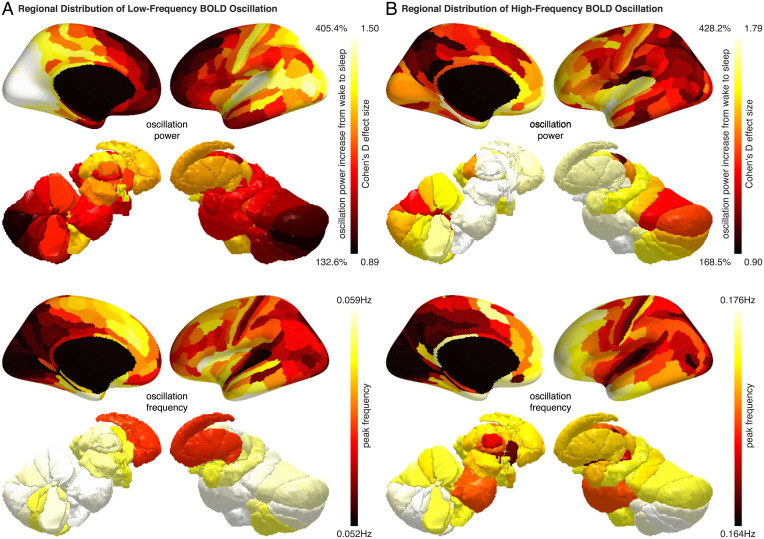
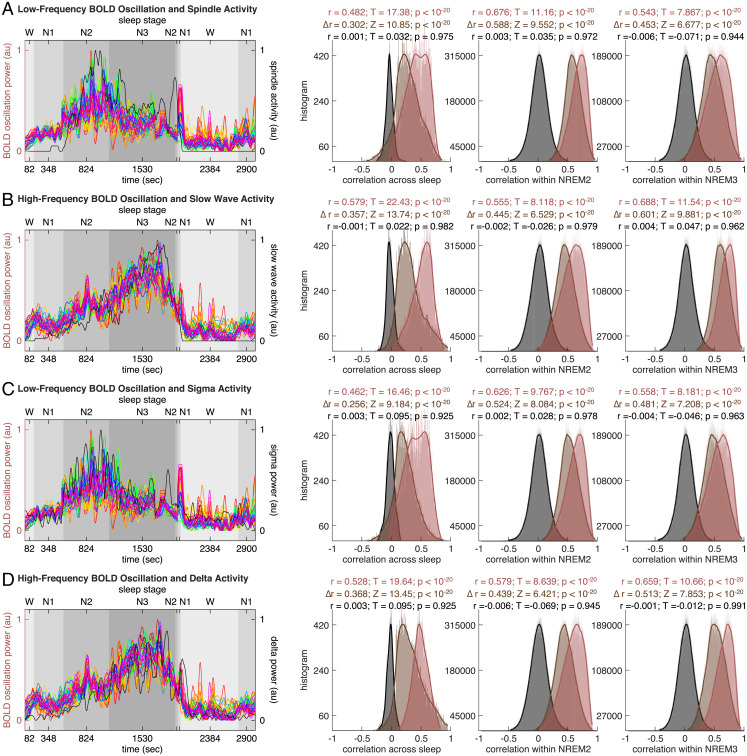
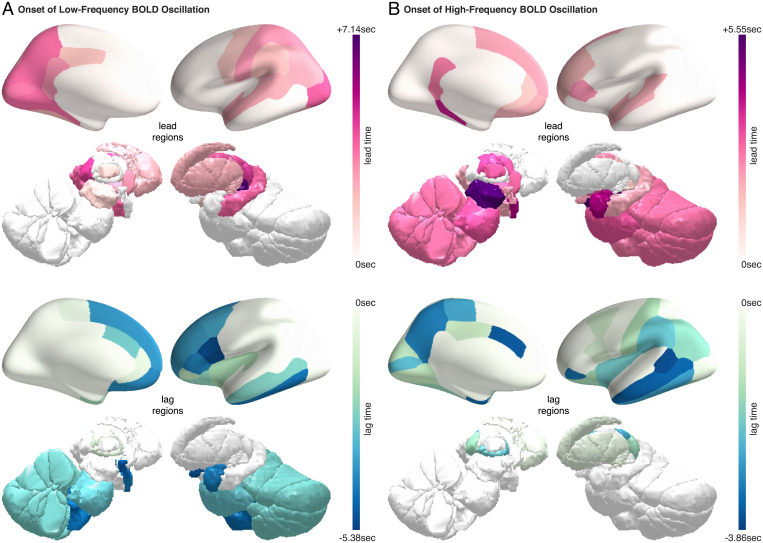
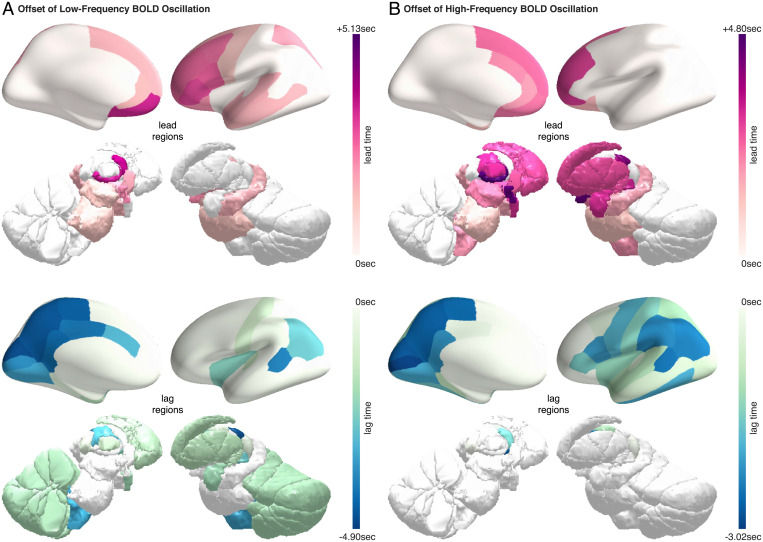
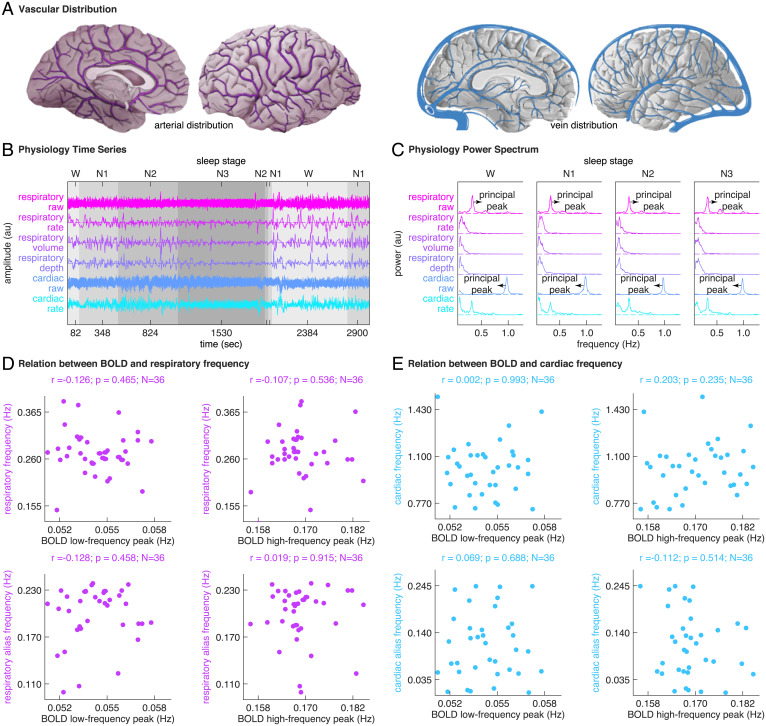
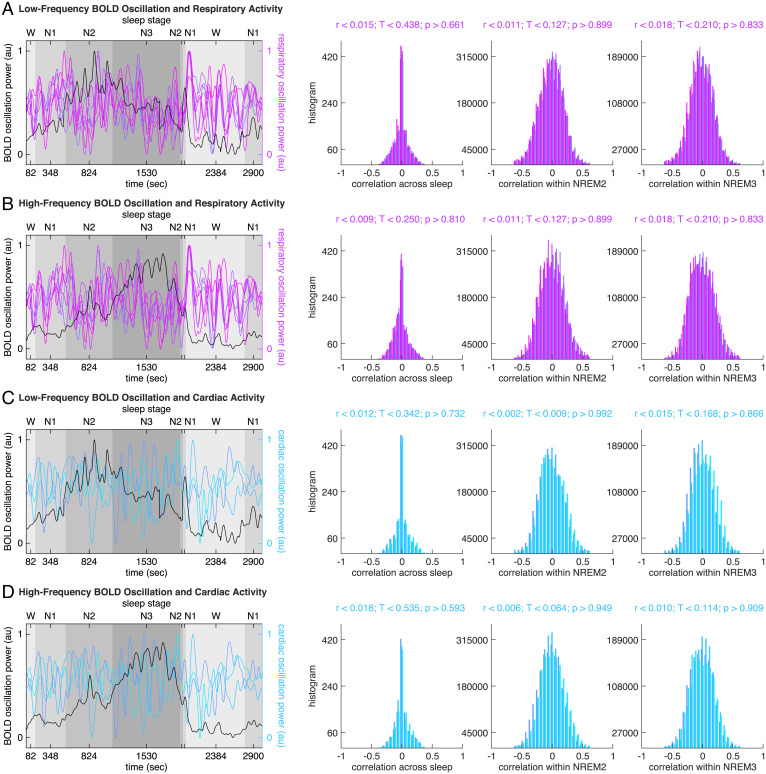
Sleep can be distinguished from wake by changes in brain electrical activity, typically assessed using electroencephalography (EEG). The hallmark of nonrapid-eye-movement (NREM) sleep is the shift from high-frequency, low-amplitude wake EEG to low-frequency, high-amplitude sleep EEG dominated by spindles and slow waves. Here we identified signatures of sleep in brain hemodynamic activity, using simultaneous functional MRI (fMRI) and EEG. We found that, at the transition from wake to sleep, fMRI blood oxygen level-dependent (BOLD) activity evolved from a mixed-frequency pattern to one dominated by two distinct oscillations: a low-frequency (<0.1 Hz) oscillation prominent in light sleep and correlated with the occurrence of spindles, and a high-frequency oscillation (>0.1 Hz) prominent in deep sleep and correlated with the occurrence of slow waves. The two oscillations were both detectable across the brain but exhibited distinct spatiotemporal patterns. During the falling-asleep process, the low-frequency oscillation first appeared in the thalamus, then the posterior cortex, and lastly the frontal cortex, while the high-frequency oscillation first appeared in the midbrain, then the frontal cortex, and lastly the posterior cortex. During the waking-up process, both oscillations disappeared first from the thalamus, then the frontal cortex, and lastly the posterior cortex. The BOLD oscillations provide local signatures of spindle and slow wave activity. They may be employed to monitor the regional occurrence of sleep or wakefulness, track which regions are the first to fall asleep or wake up at the wake-sleep transitions, and investigate local homeostatic sleep processes.
Keywords: BOLD oscillations; fMRI-EEG; sleep; wake–sleep transitions.
Conflict of interest statement
The authors declare no competing interest.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Steriade M., Timofeev I., Grenier F., Natural waking and sleep states: A view from inside neocortical neurons. J. Neurophysiol. 85, 1969–1985 (2001). - PubMed
-
- Crunelli V., David F., Lőrincz M. L., Hughes S. W., The thalamocortical network as a single slow wave-generating unit. Curr. Opin. Neurobiol. 31, 72–80 (2015). - PubMed
-
- Nobili L., et al. , Dissociated wake-like and sleep-like electro-cortical activity during sleep. Neuroimage 58, 612–619 (2011). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical