Memory-enhancing properties of sleep depend on the oscillatory amplitude of norepinephrine
- PMID: 35798980
- PMCID: PMC9817483
- DOI: 10.1038/s41593-022-01102-9
Memory-enhancing properties of sleep depend on the oscillatory amplitude of norepinephrine
Abstract
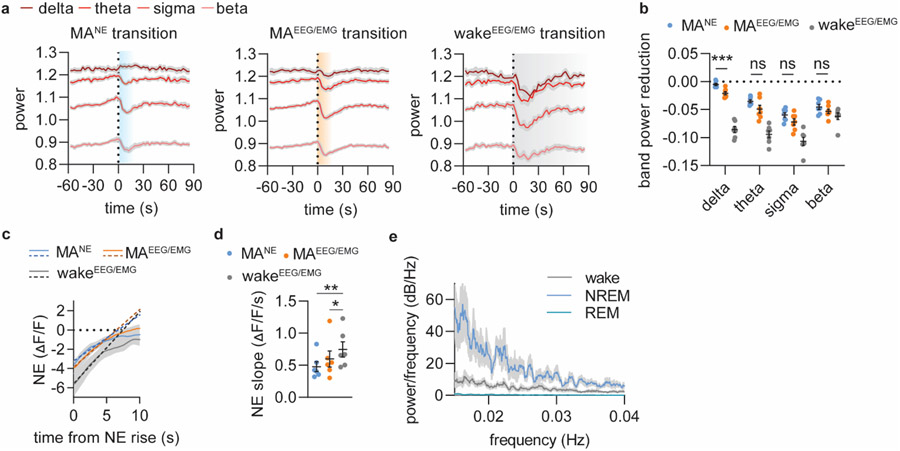
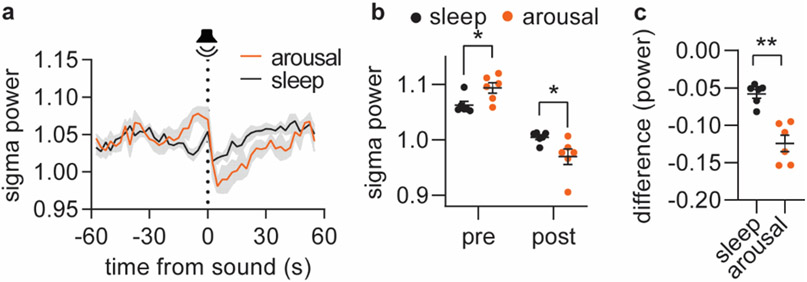
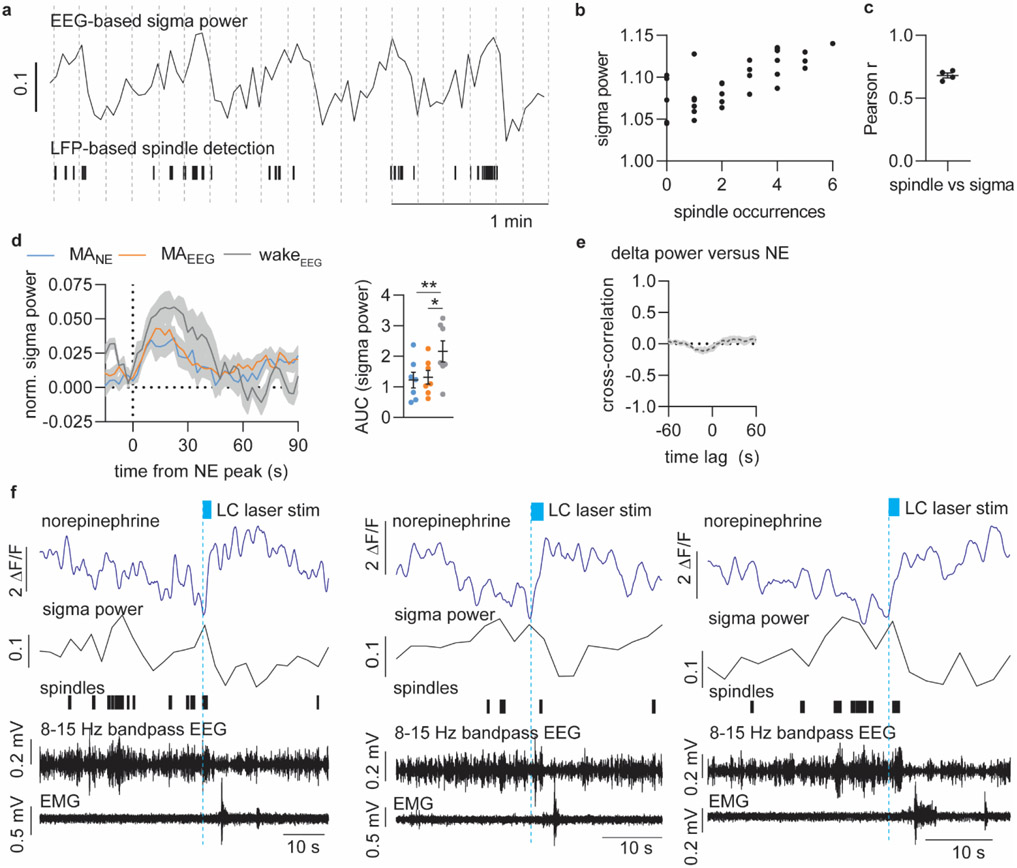
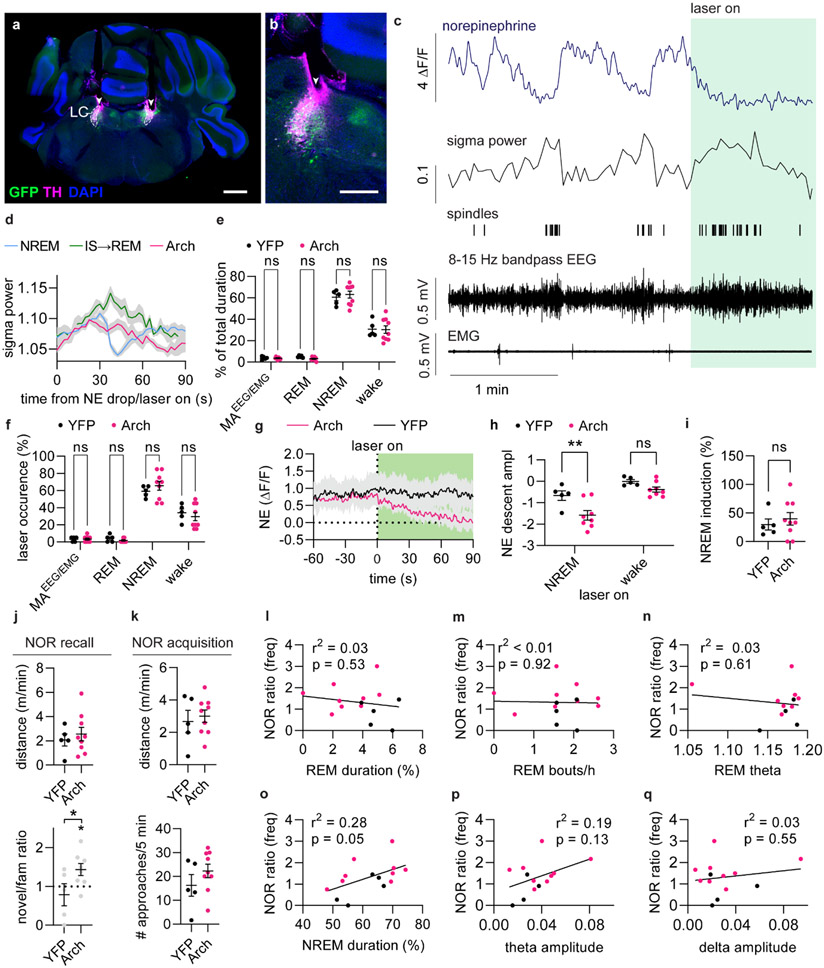
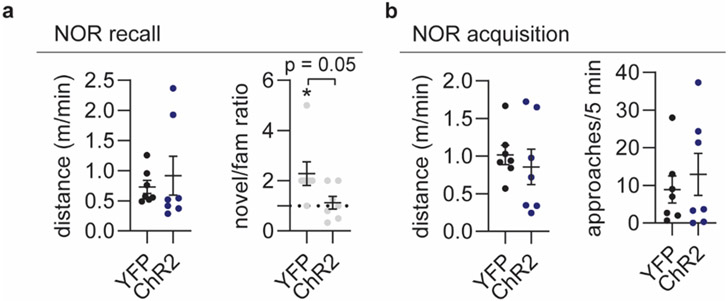
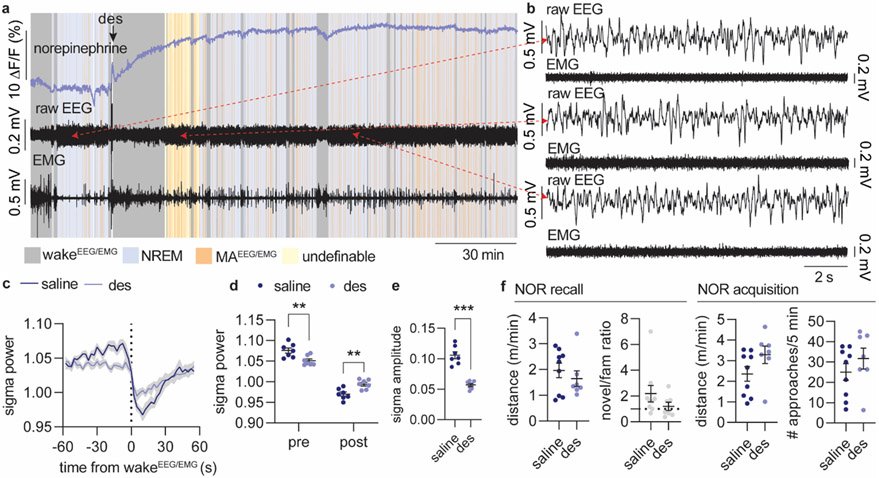
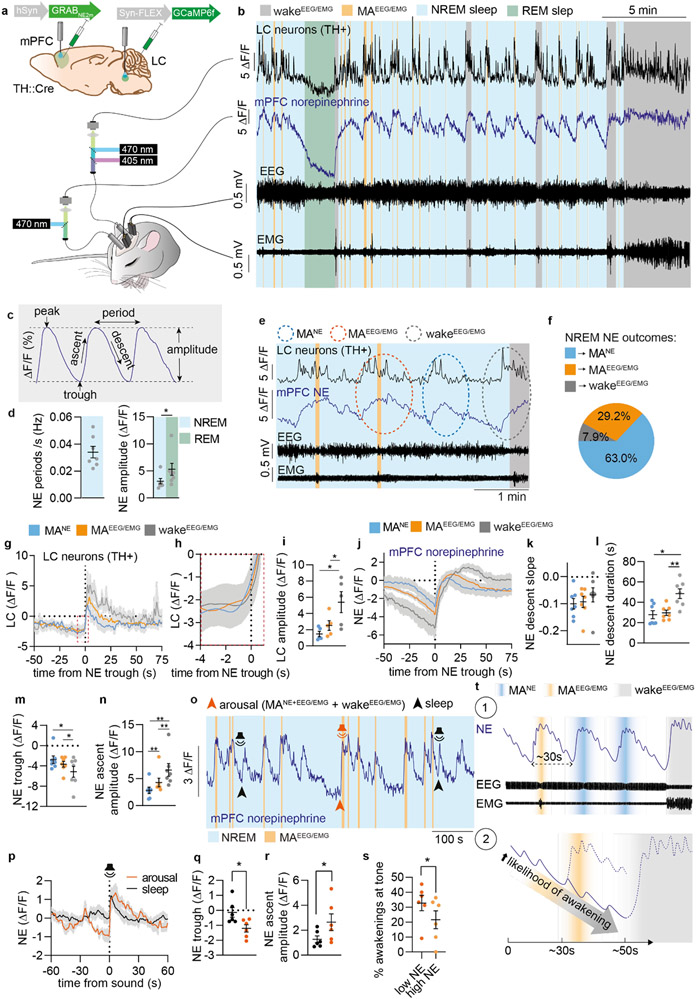
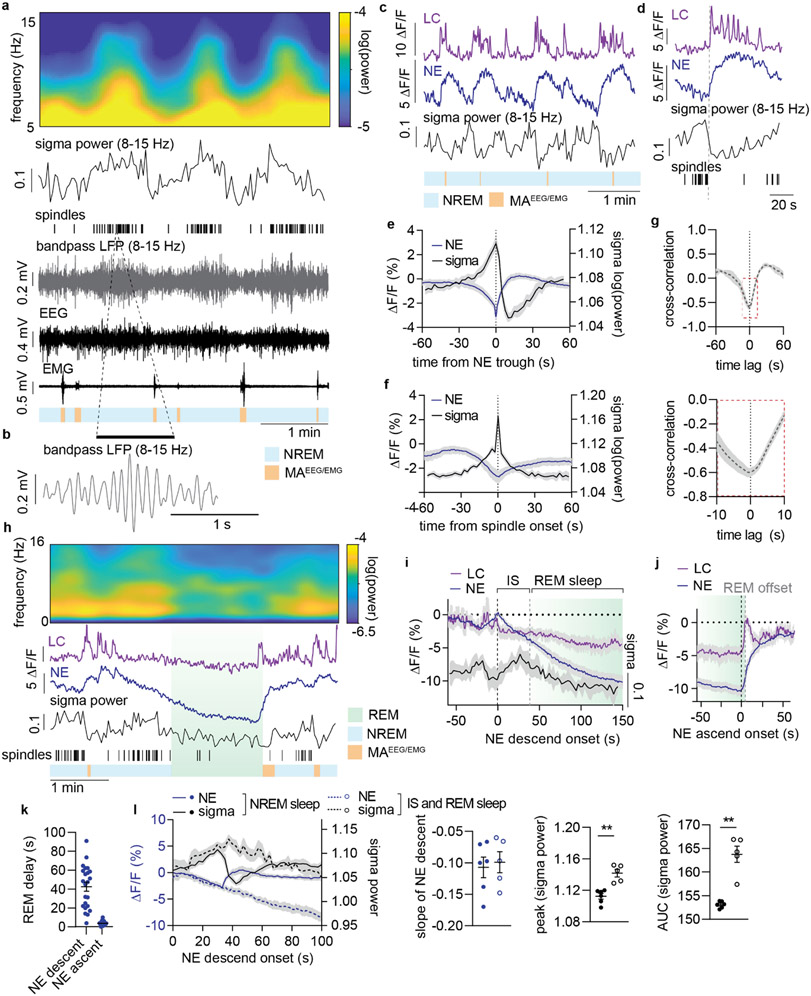
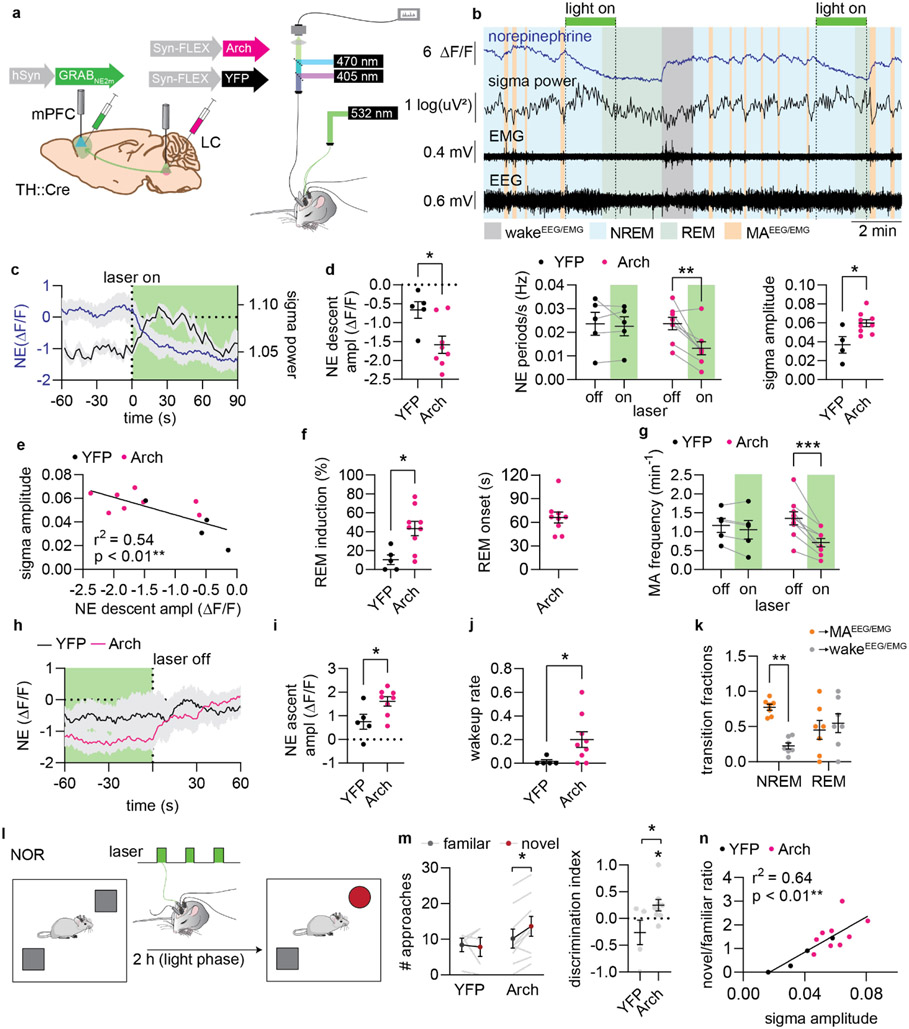
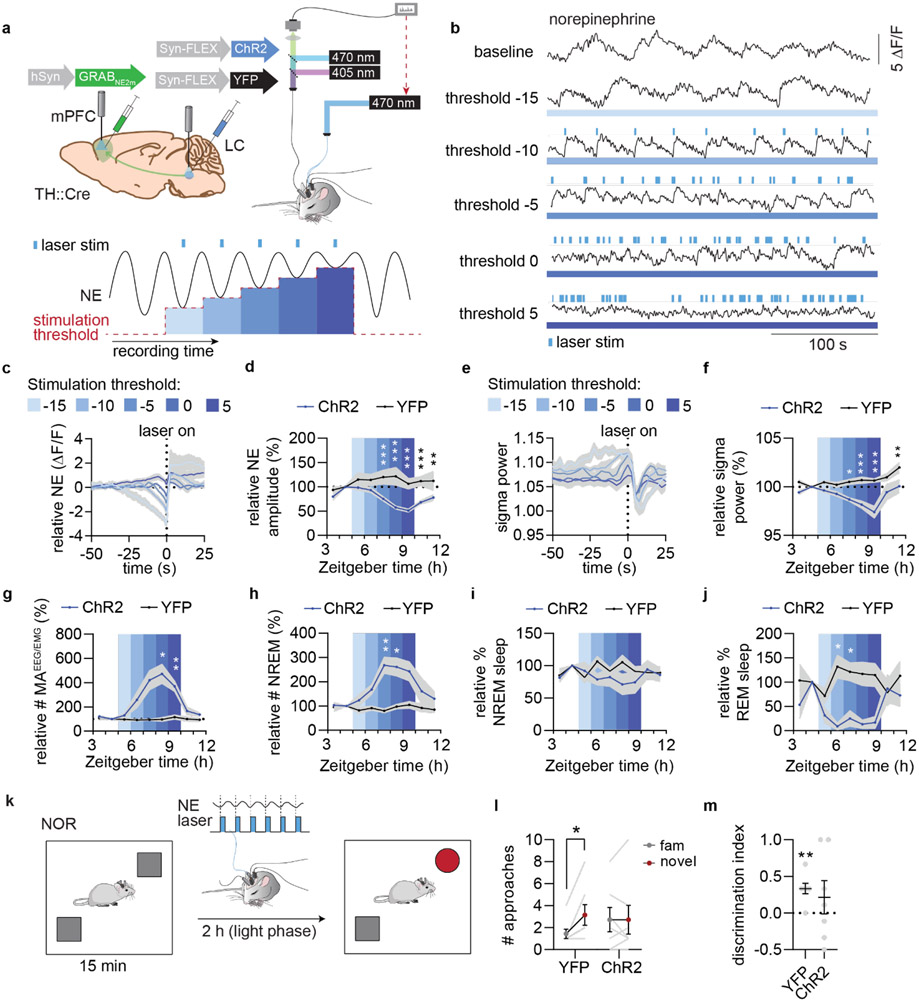
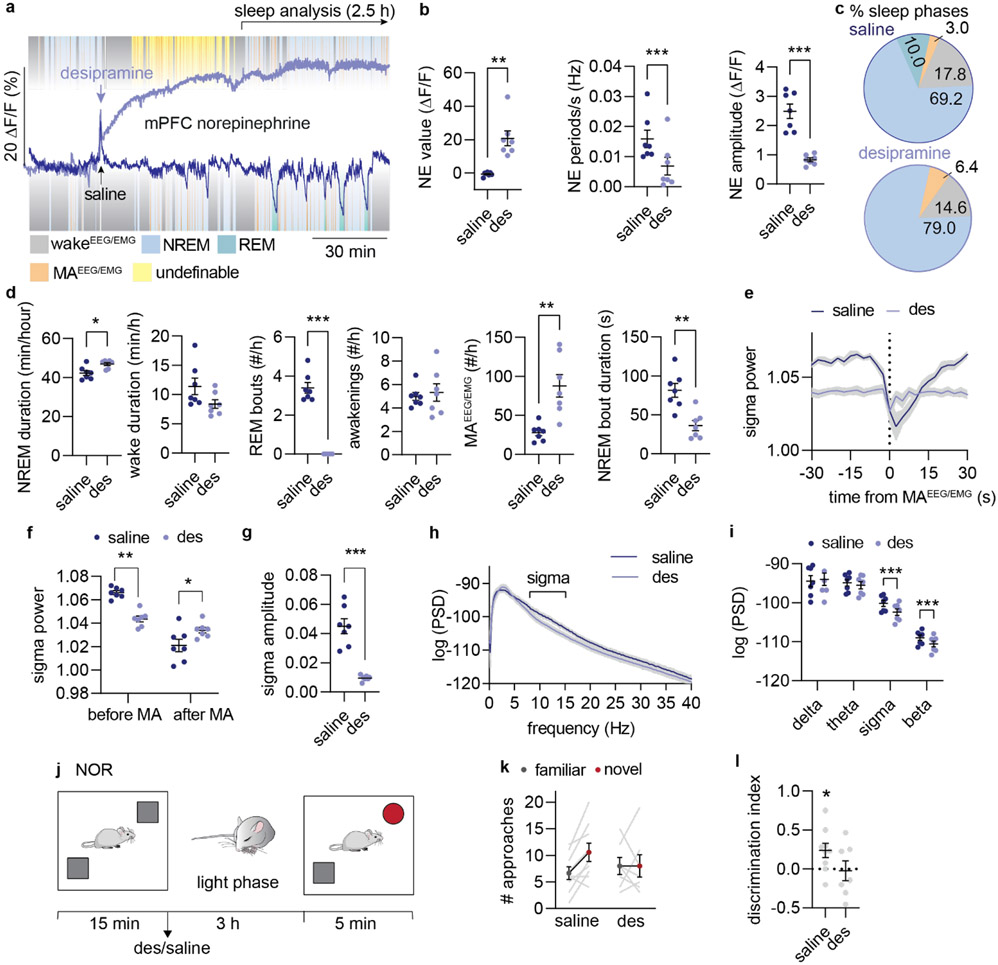
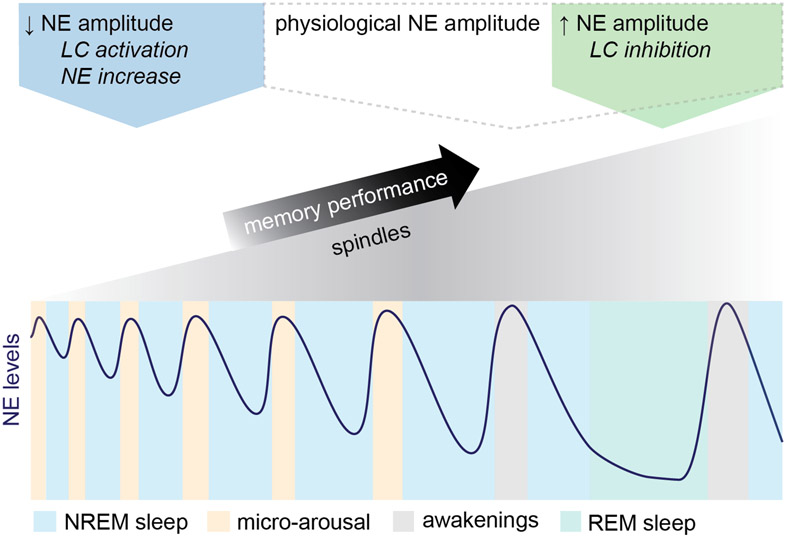
Sleep has a complex micro-architecture, encompassing micro-arousals, sleep spindles and transitions between sleep stages. Fragmented sleep impairs memory consolidation, whereas spindle-rich and delta-rich non-rapid eye movement (NREM) sleep and rapid eye movement (REM) sleep promote it. However, the relationship between micro-arousals and memory-promoting aspects of sleep remains unclear. In this study, we used fiber photometry in mice to examine how release of the arousal mediator norepinephrine (NE) shapes sleep micro-architecture. Here we show that micro-arousals are generated in a periodic pattern during NREM sleep, riding on the peak of locus-coeruleus-generated infraslow oscillations of extracellular NE, whereas descending phases of NE oscillations drive spindles. The amplitude of NE oscillations is crucial for shaping sleep micro-architecture related to memory performance: prolonged descent of NE promotes spindle-enriched intermediate state and REM sleep but also associates with awakenings, whereas shorter NE descents uphold NREM sleep and micro-arousals. Thus, the NE oscillatory amplitude may be a target for improving sleep in sleep disorders.
© 2022. The Author(s), under exclusive licence to Springer Nature America, Inc.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Comment in
-
Cortical norepinephrine GRABs a seat at the sleep table.Morici JF, Girardeau G. Morici JF, et al. Nat Neurosci. 2022 Aug;25(8):978-980. doi: 10.1038/s41593-022-01117-2. Nat Neurosci. 2022. PMID: 35798981 No abstract available.
-
Do all norepinephrine surges disrupt sleep?Lüthi A, Franken P, Fulda S, Siclari F, Van Someren EJW. Lüthi A, et al. Nat Neurosci. 2023 Jun;26(6):955-956. doi: 10.1038/s41593-023-01313-8. Epub 2023 Apr 20. Nat Neurosci. 2023. PMID: 37081297 No abstract available.
-
Reply to: 'Do all norepinephrine surges disrupt sleep?'.Kjaerby C, Andersen M, Hauglund N, Untiet V, Dall C, Ding F, Hirase H, Nedergaard M. Kjaerby C, et al. Nat Neurosci. 2023 Jun;26(6):957-958. doi: 10.1038/s41593-023-01314-7. Epub 2023 Apr 20. Nat Neurosci. 2023. PMID: 37081298 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases