Circuit mechanisms for the chemical modulation of cortex-wide network interactions and behavioral variability
- PMID: 34272245
- PMCID: PMC8284895
- DOI: 10.1126/sciadv.abf5620
Circuit mechanisms for the chemical modulation of cortex-wide network interactions and behavioral variability
Abstract
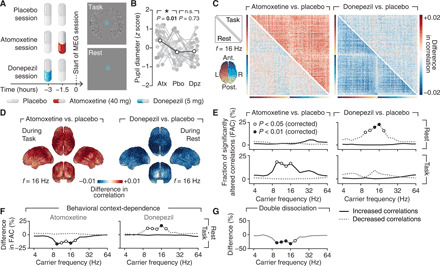
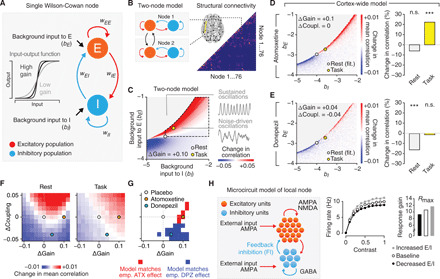
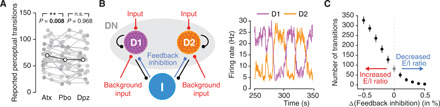
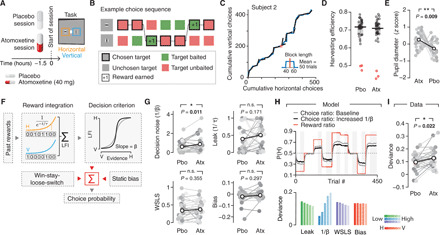
Influential theories postulate distinct roles of catecholamines and acetylcholine in cognition and behavior. However, previous physiological work reported similar effects of these neuromodulators on the response properties (specifically, the gain) of individual cortical neurons. Here, we show a double dissociation between the effects of catecholamines and acetylcholine at the level of large-scale interactions between cortical areas in humans. A pharmacological boost of catecholamine levels increased cortex-wide interactions during a visual task, but not rest. An acetylcholine boost decreased interactions during rest, but not task. Cortical circuit modeling explained this dissociation by differential changes in two circuit properties: the local excitation-inhibition balance (more strongly increased by catecholamines) and intracortical transmission (more strongly reduced by acetylcholine). The inferred catecholaminergic mechanism also predicted noisier decision-making, which we confirmed for both perceptual and value-based choice behavior. Our work highlights specific circuit mechanisms for shaping cortical network interactions and behavioral variability by key neuromodulatory systems.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Aston-Jones G., Cohen J. D., An integrative theory of locus coeruleus-norepinephrine function: Adaptive gain and optimal performance. Annu. Rev. Neurosci. 28, 403–450 (2005). - PubMed
-
- Schwarz L. A., Luo L., Organization of the locus coeruleus-norepinephrine system. Curr. Biol. 25, R1051–R1056 (2015). - PubMed
LinkOut - more resources
Full Text Sources