Combinatorial CRISPR screen identifies fitness effects of gene paralogues
- PMID: 33637726
- PMCID: PMC7910459
- DOI: 10.1038/s41467-021-21478-9
Combinatorial CRISPR screen identifies fitness effects of gene paralogues
Abstract
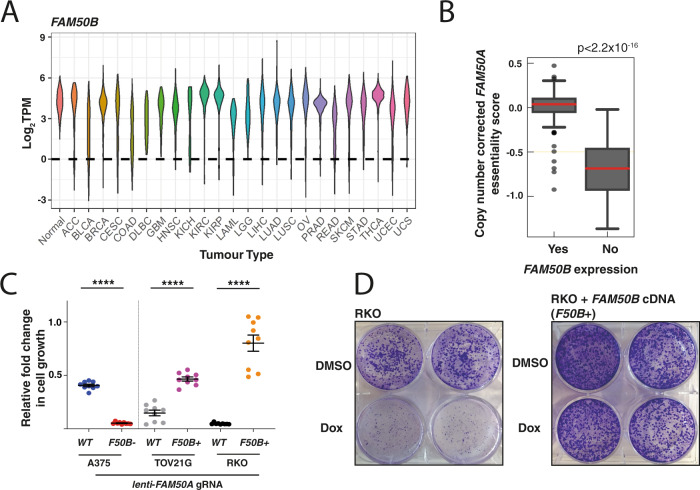
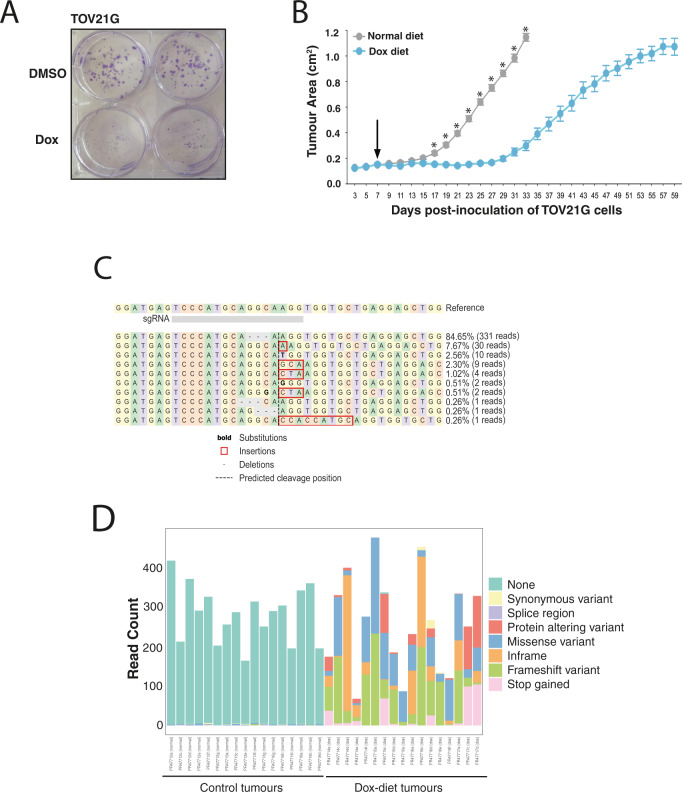
Genetic redundancy has evolved as a way for human cells to survive the loss of genes that are single copy and essential in other organisms, but also allows tumours to survive despite having highly rearranged genomes. In this study we CRISPR screen 1191 gene pairs, including paralogues and known and predicted synthetic lethal interactions to identify 105 gene combinations whose co-disruption results in a loss of cellular fitness. 27 pairs influence fitness across multiple cell lines including the paralogues FAM50A/FAM50B, two genes of unknown function. Silencing of FAM50B occurs across a range of tumour types and in this context disruption of FAM50A reduces cellular fitness whilst promoting micronucleus formation and extensive perturbation of transcriptional programmes. Our studies reveal the fitness effects of FAM50A/FAM50B in cancer cells.
Conflict of interest statement
The authors declare no competing interests.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials